GENE LOCI IN CATS
This lists the different loci and combinations of alleles, including a few hypothetical or unconfirmed genes. In the case of the posited genes listed, there has been sufficient breeding data or study of cats with the trait(s) for the genes to be statistically likely.
CLASSIC BLACK/BROWN
This is a gene series with the order of dominance: Black/Brown > Chocolate > Cinnamon. Browning genes code for tyrosinase related protein-1 (TYRP1) , an enzyme involved in the metabolic pathway for eumelanin pigment production. The dominant form, B, produces black color. Recessive variants are b (chocolate) and bl (cinnamon/light brown).
|
B - Black/Brown Locus |
||
|
Genotype |
Description |
Phenotype |
|
B/B |
Homozygous - Black/Brown |
Black/Brown |
|
B/b |
Heterozygous - Black/Brown - carrying Chocolate |
Black |
|
B/bl |
Heterozygous - Black/Brown - carrying Cinnamon |
Black |
|
b/b |
Homozygous - Chocolate |
Chocolate |
|
bl/bl |
Homozygous - Cinnamon |
Cinnamon - reddish chocolate |
|
b/bl |
Heterozygous - Chocolate - carrying Cinnamon |
Chocolate |
"BARRINGTON BROWN"
Don Shaw's "Barrington Brown" may be a further recessive brown colour. It was observed in a colony of laboratory cats, none of which left the laboratory. Some breeders erroneously believed it to have been the dilute modifier gene (which turns eumelanin-based colours to caramel). However, the Barrington Brown homozygotes were different in colour to caramel cats. In the homozygous form, Barrington Brown had an additive effect on the other colours (some suggest cats with one copy of the gene and cats with two copies looked different, but a cat with only one copy of the gene shouldn't show any effect).
|
ba - Barrington Brown Locus (unverified) |
||
|
Genotype |
Description |
Phenotype |
|
Ba/Ba |
Homozygous - Non Barrington Brown |
Cat unaffected - i.e. Black/Brown/Chocolate etc |
|
Ba/ba |
Heterozygous - Non Barrington Brown - carrying Barrington Brown |
Cat unaffected - i.e. Black/Brown/Chocolate etc |
|
ba/ba |
Homozygous - Barrington Brown -liberty of renaming |
Mahogany Brown/Light Brown/milk coffee in colour |
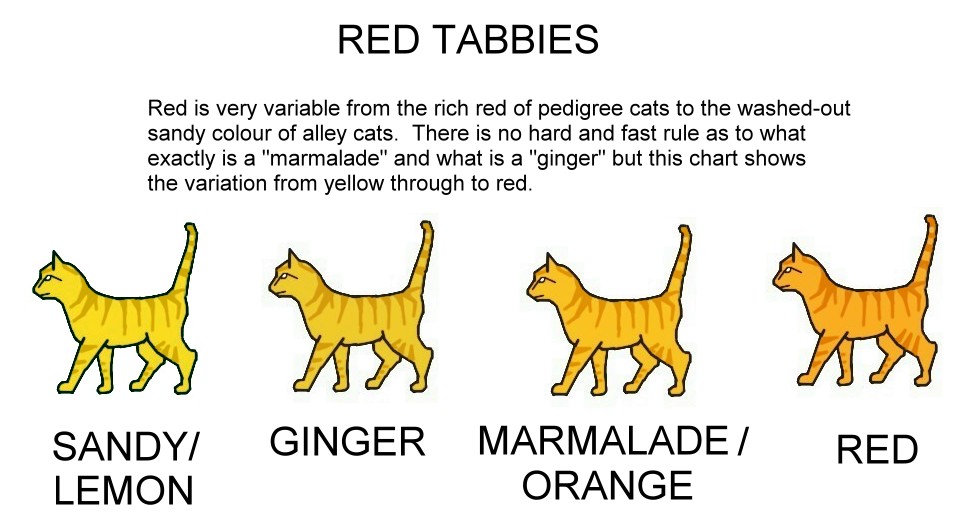
EPISTATIC RED
This is a sex-linked trait carried on the X chromosome. It causes phaeomelanin (orange pigment) to completely replace
eumelanin (black or brown pigment). Red can also be modified in the following fashion:
O + d/d (homozygous for dilute colour) = Cream
O + d/d + Dm (dilute modifier) = Apricot
|
O - Red Locus (sex-linked epistatic) |
||
|
Genotype |
Description |
Phenotype |
|
XO/XO |
Homozygous - Red Female |
Red female |
|
XO/Xo |
Heterozygous - Tortie |
Female, Red intermixed with a non-red colour i.e.. black tortie, chocolate tortie |
|
Xo/Xo |
Homozygous - Non-red female |
Female, not red, not tortie |
|
XO/Y |
Red Male |
Red male |
|
XoY |
Non-red male |
Male, not red |
|
XXY |
Klinefelter mosaic male |
Sterile tortie male (red + a non-red colour) |
|
XY/XY, XX/XY |
Chimera mosaic male |
Fertile tortie male (red + a non-red colour), passes on either red or non-red to offspring |
COLOURPOINT OR HIMALAYAN PATTERN (ALBINO SERIES)
Colourpoint is a series of alleles (also known as temperature-sensitive albino) due to 2 mutations in the Tyrosinase (TYR) gene. The order of dominance is C > cb/cs > ca > c. The alleles cs and cb are co-dominant to each other and expression of these is temperature sensitive with the pigmentation being restricted to the cooler extremities of the body: muzzle, ears, tail and legs. Colourpoint cats in colder regions generally have darker body colour and more extensive points while those in warmer regions generally have paler bodies and smaller coloured areas. This can also be demonstrated by bandaging a cat's limb while it is moulting - the newly grown fur will be paler because the bandaged area is relatively warm. Colourpoint expression is also age-related, with older cats tending to be darker than younger cats.
True albino cats (absence of pigment) are rare because the more dominant alleles must be absent in order for albino to show up. Almost all blue-eyed white cats are due to the epistatic white / white spotting gene which masks pigment.
The C gene codes for the enzyme tyrosinase, the first step in pigment production.
|
C - Colour Restriction Locus |
||
|
Genotype |
Description |
Phenotype |
|
C/C |
Homozygous - Solid Coloured |
Solid colour |
|
C/c |
Heterozygous - Solid Coloured carrying red eyed albino |
Solid colour |
|
C/ca |
Heterozygous -Solid Coloured carrying blue eyed albino |
Solid colour |
|
C/cs |
Heterozygous - Solid Coloured carrying colour point |
Solid colour |
|
C/cm |
Heterozygous - Solid Coloured carrying mocha |
Solid colour |
|
C/cb |
Heterozygous - Solid Coloured carrying sepia |
Solid colour |
|
c/c |
Homozygous - Pink eyed Albino (recessive white) |
Unpigmented (white) with Pink eyes |
|
ca/ca |
Homozygous - Blue eyed Albino (recessive white) |
Unpigmented (white) with pale blue eyes |
|
cs/cs |
Homozygous - Colourpoint |
Colourpoint, Siamese pattern |
|
cm/cm |
Homozygous - Mocha |
Mocha pattern |
|
cb/cb |
Homozygous- Sepia |
Sepia, Burmese pattern |
|
cb/cs |
Heterozygous- Mink (co-dominant Sepia and Colourpoint) |
Mink, Tonkinese pattern (intermediate between colourpoint and sepia) |
|
cb/cm |
Heterozygous- Burmocha (co-dominant Sepia and Mocha) |
Intermediate between Burmese sepia and Mocha pattern |
|
cs/cm |
Heterozygous- Siamocha (co-dominant Colourpoint and Mocha) |
Intermediate between Colourpoint and Mocha pattern |
|
cs/ca |
Heterozygous - Bondanese |
Colourpoint, Siamese with high contrast |
|
cb/ca |
Heterozygous- Sepia & Blue eyed albino |
Sepia, Burmese pattern (unverified) |
|
cs/c |
Heterozygous Point & Red eyed albino |
Sepia, Burmese pattern (unverified) |
|
cb/c |
Heterozygous Sepia & Red eyed albino |
Sepia, Burmese pattern (unverified) |
|
ca/c |
Heterozygous Blue eyed albino & Red eyed Albino |
Unpigmented (white) with pale blue eyes (unverified) |
Breakthrough Tabby Pattern in Colourpoints
Cats born of Siamese parents sometimes have a tabby/spotted pattern breaking through so strongly that the cat appears to be a tabby/spotted Oriental or a tabby/spotted sepia (Burmese) cat. Viewed closely, the pattern colour is heavily ticked on a paler ticked background. This breakthrough pattern is evident from kittenhood and not the same as age-related darkening in Colourpoint cats. It is inherited, possibly due to modifier genes that prevent the colourpoint gene from properly inhibiting the colour/pattern on the normally pale areas of the body.
COLOUR DILUTION & DILUTE MODIFIER
Dense and Dilute colours are due to a simple gene pair (gene: Melanophilin or MLPH). When two dilute alleles are present, it changes the basic (dense) colour to its dilute form e.g. black dilutes to blue, chocolate dilutes to lilac, cinnamon dilutes to fawn, red dilutes to cream. The dominant Dilute Modifier gene only affects dilute colours. Dilute modifier has no effect on Dense colours, but may be carried by dense-colour cats and passed on to offspring and may therefore show up in later generations that also have the two dilute alleles.
D/d, codes for melanophilin, a protein involved in the transportation and deposition of pigment into a growing hair. The dominant form allows denser deposition of pigment.
|
D - Colour Density/Dilution Locus |
||
|
Genotype |
Description |
Phenotype |
|
D/D |
Homozygous - Dense pigmentation |
Dense (dark) colour, e.g. black, chocolate, cinnamon, red |
|
D/d |
Heterozygous - Dense carrying dilution |
Dense (dark) colour, e.g. black, chocolate, cinnamon, red |
|
d/d |
Heterozygous - Dilute |
Dilute colour, e.g. blue, lilac, cinnamon, cream |
|
Dm - Dilute Modifier Locus |
||
|
Genotype |
Description |
Phenotype |
|
Dm/Dm |
Homozygous - Dilute Modifier |
Modified dilute colour (turns dilute eumelanin colours to caramel, turns cream to apricot), no effect on dense colour |
|
Dm/dm |
Heterozygous - Dilute Modifier - carrier of non dilute modifier |
Modified dilute colour (turns dilute eumelanin colours to caramel, turns cream to apricot), no effect on dense colour |
|
dm/dm |
Homozygous - Non-dilute Modifier |
No effect |
EPISTATIC (DOMINANT) WHITE, WHITE SPOTTING, MITTING AND GLOVING GENE
Research has found that White Masking and White Spotting are dominant alleles of the same gene (called KIT) and the order of dominance is W >> Ws >> w >> wg.
W produces solid white and is the most dominant in the series. Ws is extremely variable in how it shows up; it is dominant over solid colour (non-white-spotted). A cat with a single copy of Ws might have a small amount of white or a large amount. Most often, though, cats with two copies of Ws have more white, and may even be solid white. Selective breeding can fix the amount of white because other genes, known as polygenes, can influence colour distribution e.g. Some breeds have been fixed for gloves or mitts. A separate recessive gene wg has been identified for Birman white gloving. There are hypothetical genes for other white mitted patterns and for the York Chocolate white pattern. There are other genes, as yet unidentified, for the white throat locket and white brisket spots.
White prevents migration of pigment-producing cells to the skin during embryologic development. The number of melanocytes are greatly reduced, but not always absent (hence temporary smudges of colour on the heads of some kittens). Dominant white ( W ) is associated with developmental defects where melanocytes fail to migrate properly. This can result in one or both blue eyes and in degenerative changes in the succule and cochlea of the ear (resulting in partial or total deafness). White spotting ( Ws ) is variable in expression from white locket through to solid white, and is rarely associated with deafness.
The Birman "White Gloving" pattern ( wg previously called G ) is the most recessive of the alleles identified. A cat with two copies of wg has gloved white feet and this trait has been fixed other white markings bred out - so that Birmans are homozygous for wg. Cats with the White Glove mutation are also found in the Ragdoll, Egyptian Mau, Exotic Shorthair, Maine Coon, Manx, Seychellois, Siamese, Siberian, Sphynx and Turkish Van. Testing has found that the Ragdoll "Mitted" pattern is different from wg. Because white feet can result from several different mutations, a genetic test has been developed for the White Gloving gene.
|
W - Dominant White / White Spotting Locus |
||
|
Genotype |
Description |
Phenotype |
|
WW |
Homozygous white |
Solid white |
|
WWs |
Heterozygous white carrying white spotting |
Solid white |
|
Ww |
Heterozygous white carrying non-white |
Solid White |
|
Wwg |
Heterozygous white carrying white gloving |
Solid white |
|
Wwsal |
Heterozygous white carrying salmiak pattern |
Solid white |
|
WsWs |
Homozygous white spotting |
Variable amount of white spotting / bicolour. |
|
Wsw |
Heterozygous white spotting carrying non-white |
Variable amount of white spotting / bicolour. |
|
Wswg |
Heterozygous white spotting carrying white gloving |
Variable amount of white spotting / bicolour. |
|
ww |
Homozygous non-white |
No white markings |
|
wwg |
Heterozygous non-white carrying white gloving. |
No white markings |
|
wgwg |
Homozygous white gloving |
White gloving (note: there may be other gloving/mitting genes) |
|
wsalwsal |
Homozygous salmiak |
Salmiak pattern (exact interaction with other white spotting genes unclear) |
|
Sb or Sm - White Mittening Locus (posited - non-Birmans) |
||
|
Genotype |
Description |
Phenotype |
|
Sb/Sb or Sm/Sm |
Homozygous - Incompete dominance - white mitted |
White mitts with variable expression. |
|
Sb/sb or Sm/sm |
Heterozygous - White mitted carrying non-mitted |
White mitts with variable expression. |
|
sb/sb or sm/sm |
Homozygous - Non mitted |
No white mitts. |
|
Spy - York Chocolate White Patterning Locus (posited) |
||
|
Genotype |
Description |
Phenotype |
|
Spy/Spy |
Homozygous - Consistent white pattern |
Consistent York Chocolate white pattern |
|
Spy/spy |
Heterozygous - Consistent white pattern carrying non-white-patterned |
Consistent York Chocolate white pattern |
|
spy/spy |
Homozygous - Non-white-patterned |
No white pattern |
|
For reference, here is the older notation you will find in some books:-
|
|||||||||||||||||||||||||||||||||||||||
TABBY AND TABBY MODIFIERS
The agouti gene (A/a) determines whether the background colour is agouti or solid i.e. whether the tabby genes present are expressed or not. It codes for agouti signalling protein (ASIP). The dominant, wild-type A causes the agouti shift phenomenon which causes hairs to be black pigmented at the tips and orange pigmented at the roots, allowing any underlying tabby pattern to be revealed. The recessive non-agouti (or hypermelanism) allele, prevents this shift in the pigmentation pathway and hides the tabby pattern (apart from ghost striping). The sex-linked O (orange) gene is epistatic (dominant) over non-agouti, hence red cats still have tabby markings (particularly visible in tortie cats where solid black patches and red-tabby are interspersed).
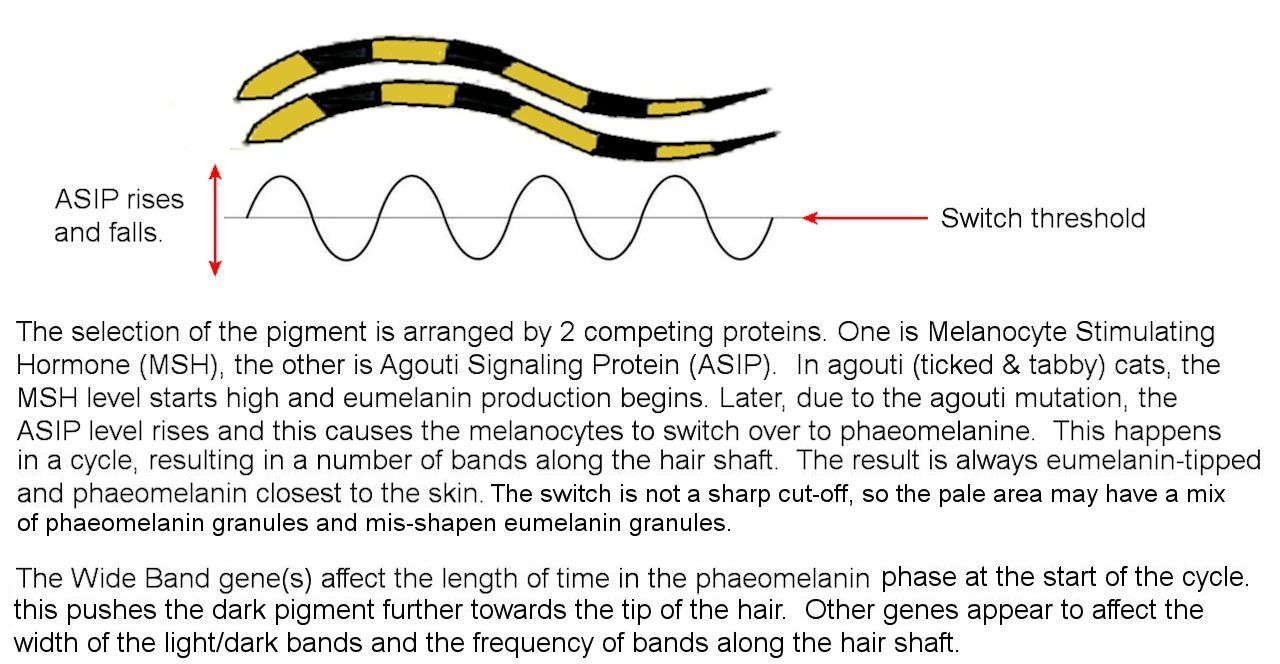
The secondary tabby locus is also called patterned/unpatterned tabby and determines whether an underlying blotched or striped pattern is manifested or not (in essence, it givesor denies permission for the tabby pattern to be expressed). The primary tabby locus determines the tabby type - classic or mackerel. The spotted pattern modifier changes these two patterns into spotted tabbies.
There are other hypothetical modifier genes affecting the overall pattern: Chaos (intermixes dark and light areas, disrupting the normal striped pattern), Confusion (adjacent hairs have different numbers and widths of colour bands) , Unconfused (adjacent hairs have consistent numbers and widths of colour bands), Erase (erases residual markings on limbs), and Roan (intermixes solid white hairs with coloured hairs). Tabby cats with both a colour inhibitor (silver) gene and the Wide Band gene will appear as shaded or tipped cats while those lacking Wide Band will be silver tabbies.
An unidentified "grizzled" gene in Chausie cats, that may be related to agouti gene, produces silver-tipped black fur and appears to have been inherited from the breed's wild Jungle Cat ancestors in which this trait is found.
|
A - Agouti Locus |
||
|
Genotype |
Description |
Phenotype |
|
A/A |
Homozygous - Agouti |
Banding present on the hair shaft, tabby |
|
A/a |
Heterozygous - Agouti - carrying non-agouti |
Banding present on the hair shaft, tabby |
|
a/a |
Homozygous - Non-Agouti |
No banding of the hair shaft, solid |
|
Ta - Secondary (Ticked) Tabby Locus |
||
|
Genotype |
Description |
Phenotype |
|
Ta/Ta |
Homozygous - Unpatterned tabby |
Ticked (Abyssinian-type) tabby, no pattern (some residual barring on lags, tail and face) |
|
Ta/ta |
Heterozygous - Agouti - carrying non-agouti |
Ticked (Abyssinian-type) tabby, no pattern (some residual barring on lags, tail and face) |
|
ta/ta |
Homozygous - Non-Agouti |
No effect. Mackerel or Classic tabby depending on which Primary Tabby gene is present and whether the cat is agouti or non-agouti. |
Mackerel and Classic tabbies are modified by the ticked tabby allele and the spotted tabby allele. They must have non ticked (a/a) and non-spotted (sp/sp) alleles both present for the Mackerel or Classic tabby pattern to show up.
Spotted Tabby is a dominant modifier of both Mackerel and Classic Tabby, but is hypostatic to Ticked Tabby (is masked by epistatic ticked tabby gene). It modifies both tabby patterns into spotted patterns.
|
Mc - Primary Tabby Locus |
||
|
Genotype |
Description |
Phenotype |
|
Mc/Mc |
Homozygous - Mackerel Tabby |
Mackerel (Tiger) Tabby |
|
Mc/mc |
Heterozygous- Mackerel - carrying Classic Tabby |
Mackerel (Tiger) Tabby |
|
mc/mc |
Homozygous - Classic Tabby |
Classic (Blotched) Tabby |
|
Sp - Spotted Tabby Locus |
||
|
Genotype |
Description |
Phenotype |
|
Sp/Sp |
Homozygous - Spotted Tabby |
Spotted Tabby |
|
Sp/sp |
Heterozygous - Spotted Tabby - carrying non spotted |
Spotted Tabby |
|
sp/sp |
Homozygous - Non Spotted Tabby |
No effect |
Breeder observations and analysis of pedigrees have resulted in several posited tabby modifiers. The Tabby Pattern Size modifier modifies the Classic Tabby Pattern by creating pale areas within the blotched markings and may be the same as the hypothetical Chaos modifier.
The Tabby Pattern Spot Modifier converts the 2 basic tabby patterns into spotted patterns. Spotted mackerel tabby will have smaller, more numerous, more vertically aligned spots while spotted classic tabby will have larger spots with less evident vertical alignment.
|
Th - Tabby Pattern Size Modifier Locus (posited) |
||
|
Genotype |
Description |
Phenotype |
|
Thl/Thl |
Homozygous - unmodified classic tabby pattern |
Classic Tabby |
|
Thl/ths |
Heterozygous - classic Tabby carrying Sokoke pattern |
Classic Tabby |
|
Ths/ths |
Homozygous - small pattern classic tabby |
Sokoke-pattern classic tabby with pale areas within the dark markings. |
|
Pm - Tabby Pattern Spot Modifier Locus (posited) |
||
|
Genotype |
Description |
Phenotype |
|
Pmf/Pmf |
Homozygous - modified tabby pattern |
Spotted tabby (large spots when interacting with classic tabby, small spots when interacting with mackerel tabby) |
|
Pmf/pmu |
Heterozygous - modified tabby carrying unmodified tabby |
Spotted tabby (large spots when interacting with classic tabby, small spots when interacting with mackerel tabby) |
|
Pmu/pmu |
Homozygous - unmodified tabby pattern |
Classic tabby or mackerel tabby, non-spotted |
There is no official symbol for the Bengal Marble effect (and its "marbled" and "clouded" analogues in other spotted hybrids), but breeders sometimes use "mz". It appears recessive to the spotted/rosetted pattern in the same way that classic (blotched) tabby is recessive to mackerel tabby. Marble may be the classic tabby gene modified by wild-type genes. To indicate the mode of inheritance I've simplified spotted/rosetted as being the dominant "effect" with marble being the recessive "effect" (note I am not calling them genes as I suspect they involve interactions of domestic tabby with wild spotting genes). "Bg" is not an official gene symbol.
|
Bg - Bengal Spotted/Rosette/Marble Pattern |
||
|
Genotype |
Description |
Phenotype |
|
Bg/Bg |
Homozygous - Spotted/Rosetted |
Spotted/Rosetted |
|
Bg/bg |
Heterozygous - Spotted/Rosetted - carrying Marble (Cloud) |
Spotted/Rosetted |
|
bg/bg |
Homozygous - Marble (Cloud) Pattern |
Marble (Cloud) |
CHARCOAL (AGOUTI VARIANT)
The following information is courtesy Joshua Dabbs who is investigating charcoal Bengals with Terra Sinclair.
"Charcoal" is found in Bengals, especially in the early generations, and has also occurred in Savannahs due to Bengal outcrosses in breed development. Charcoal Bengals are "Aa" at the agouti locus. Charcoal is due to a variant in the agouti gene ("charcoal agouti" or APb variant) inherited from the Asian Leopard Cat reacting with the domestic cat non-agouti gene (a). Non-agouti genes develop independently in different species and though the visual effect is similar, the gene mutations are different. In hybrids, they interact to produce novel effects. The genotype APba results in an intermediate form between agouti and non-agouti. The intermediate form causes an increase in black pigmentation unevenly distributed throughout the coat. The distribution of pigmentation varies between cat, and often causes coloration and features to become more exaggerated to where they take on an entirely new appearance. The domestic cat version of the Agouti (A) gene is termed AFc in the table below.
Charcoal is now seen many generations down the line. Joshua Dabbs and Terra Sinclair investigated a number of pedigrees that supported that theory on inheritance of charcoal. These include a Bengal melanistic carrier that when paired with an F-Generation Bengal (F1 to F4 = 1 to 4 generations removed from a wild parent) produced charcoal colored brown spotted offspring. Early on in the breed history, charcoals were not properly identified as they they did not always pass their colouration to the next generation. They were misidentified as minks, smokes (from silver smoke Egyptian Mau outcrosses) or unrufoused browns. A high degree of rufism was sought in the original Bengals and the seemingly unrufoused browns were undesirable. A greater interest in the unrufoused Bengals (e.g. in silvers and melanistics (smokes)) has increased interest in, and acceptance of, the charcoal series.
|
A - Agouti Locus |
||
|
Genotype |
Description |
Phenotype |
|
APb/APb |
Homozygous - Agouti (ALC gene) |
High contrast Bengal pattern |
|
AFc/APb |
Heterozygous - Domestic gene agouti and ALC agouti |
Normal contrast Bengal pattern |
|
AFc/AFc |
Homozygous - Agouti (Domestic gene) |
Normal contrast Bengal pattern |
|
APb/a |
Heterozygous - Charcoal - carrying non-agouti |
Charcoal Bengal pattern |
|
a/a |
Homozygous - Non-Agouti |
No banding of the hair shaft, "melanistic" Bengal |
Charcoal Bengals vary in appearance though there are identifying factors known as "Zorro" markings. Charcoals have pure white or off-white goggles around the eyes while smokes have ghost tabby markings. Accompanying the white goggles there is usually a dark "charcoal mask" from the forehead, down the nose-bridge and often underneath the eyes and extending into the mascara-lines; the mask resembles an upside-down "Y". Charcoals also have an unbroken "cape" or "sheet" down their backs (broad back-stripe), in some cats this extends into the patterns closest to the spine-line while others have a narrower "modified" cape running down the back. Charcoal Bengals, including the browns, tend to have silver glitter, rather than either gold glitter (on normal tawny colours) or crystal glitter (on the snows and silvers).

Charcoal Brown is the most common charcoal colour; some have a uniform charcoal colour, while others have lighter background on their sides and darker background towards the top and tail. Charcoal Browns may be born looking silver due to black markings on a light greyish, background. Others ("black charcoals") are born so dark they may appear melanistic, but the background lightens as the kittens grow. Charcoal silver, when paired with a regular silver Bengal, results in higher contrast in the kittens. Charcoal silvers may be born very dark, resembling a silver smoke except for the white facial markings, becoming black-on-silver as they grow; the darker ground colour also masks the appearance of tarnishing. Colourpoint variants show darker pigmentation in their pattern or the pointing (e.g. in lynx point) is more prominent due to the higher level of pigmentation. Charcoal Snows have a greater contrast with darker markings than regular snows. Charcoal Marbles have tended to carry a sheet of colour rather than having distinct marble patterning.
Charcoal Savannahs have white or off-white goggles around the eyes and a distinctive dark nose. Rather than a pronounced mask or cape (as in bengals), there appear to be dense tabby markings between the ears. The background colour is grey rather than golden/tawny. The colour seems to trace back to the Kirembo line and has turned up in the USA and UK.
COLOUR EXTENSION: NORWEGIAN FOREST CAT "AMBER", "CARNELIAN"/"COPAL" KURILIAN BOBTAIL, "GOLDEN" SIBERIAN, BURMESE "RUSSET", BENGAL "SORREL" & "RUFOUSING"
The Extension gene (symbol E) or red factor controls the production of red and black pigment. The dominant allele E produces black pigment in the coat while the recessive allele e produces red pigment. The name comes from the effect of black or brown pigment not being extended throughout the whole coat, but being restricted to the skin of the extremities and to the eyes (for example in bay horses). Amber is due to a recessive mutation in the MC1R gene.
In Norwegian Forest Cats, this brightens black areas of the coat, modifying Black and Blue to Amber and Light Amber respectively. It has not yet been found in combination with other genes recessive to black. So far, it has only been observed in Norwegian Forest Cats where cinnamon and fawn alleles are known to be absent. It has also been called "fox". It may be present in other breeds, but its similarity to fawn and cinnamon means it may not have been identified as the Extension Gene (also known as Black Modifier or Agouti Modifier) in those breeds. The extension locus (the melanocortin receptor) or its ligands (molecules that bind to that receptor), control melanocyte stimulating hormone and agouti signaling protein.
Similar to amber is russet, which turned up in a line of seal (brown) European-style Burmese in New Zealand in 2007. It has subsequently occurred found in the related Mandalay (similar to the Asian in Europe) and appears to be a mutation of the extension gene. The first known russet was a pure-bred Burmese called Molly in 2007. There is now an experimental programme in NZ to breed Russet Burmese and to investigate dilute russet, russet tabby and solid russet (as opposed to the Burmese sepia form of russet).
The first russet kitten Molly was born an "odd-coloured lilac (lavender)" which gradually lightened as she grew, progressing through lilac-caramel and chocolate ticked tabby and then dramatically changing to red. Chocolate ticked tabby and red were both impossible from her pedigree, and in any case, reds are not born as chocolate tabbies! Several more unusually coloured kittens were born in different litters and all went through the same colour/pattern changes. The ancestors of these kittens were seal Burmese and had no silver, tabby or Mandalay (Asian) blood. The pedigree had both dilute and chocolate, there were no reds, creams or torties until 4 generations back. DNA testing showed some kittens to be genetically seal and other genetically chocolate. Hence the new colour is due to a different mutation currently known as russet. Russet appears to be due to a recessive mutation that causes black pigment (eumelanin) to gradually fade to a minimal amount while leaving red pigment (phaeomelanin)unaffected.
Russet kittens resemble tabbies at birth, but have pink noses and paw-pads, pale fur around the pads and genitalia and a pale tail-tip - all of which would be dark in tabbies. The muzzle and fur around the eye is ivory. The back is solidly dark rather than ticked, becoming pale ivory halfway down the flanks. The back becomes more ticked appearance, almost a saddle, as kittens undergo the colour change and the face becomes reddish. By age two, they may resemble a red Burmese. It differs from amber as ambers have dark noses and paw-pads. Non-agouti (solid) amber kittens are very dark with a dark face that is last to go red while russet kittens have off-white faces (possible due to Burmese sepia gene in the mix), which are the first part to go red (rather than the last as in ambers), and pale undersides. The russet colour change appears to be slower than the amber colour change. Russet kittens to date have been larger at birth than their siblings and somewhat on the large side as adults.
In the Kurilian Bobtail, a dominant colour change gene has been identified. Copal (also known as Carnelian to breeders) causes brown tabby kittens to be born with reddish hues and turn into red tabbies with a dark tail-tip in adulthood. its effect on blue or non-tabbies has not been described yet.
"Golden" or "golden tabby" Siberians have a recessive Corin mutation which is a wide band gene. The name "zoloty" (Russian for golden) was suggested for this mutation because"golden" is already used for non-silver tipped, shaded and smoke cats. It is now called Sunshine.
A similar effect is seen in Bengals where different Corin mutations cause "Sorrel" Bengals where the pattern colour has faded from black to reddish brown. Bengal breeders also believe there is a rufousing gene that gives a very reddish background without fading the markings. The background colour of early Bengals has more highly rufoused than the golden colour preferred today. They refer to the posited gene as "rf).
|
E - Extension Locus |
||
|
Genotype |
Description |
Phenotype |
|
E/E |
Homozygous - full-extension |
Allow all other colors genes to show true. |
|
E/e |
Heterozygous - full-extension - carrying non-extension |
Allow all other colors genes to show true. |
|
e/e |
Homozygous - Non-extension |
Restrict expression of eumelanin colours, convert eumelanin to phaeomelanin (primarily on torso). Black becomes Amber; Blue becomes Light Amber |
Bengal rufousing (production of red pigment in place of black/brown) is distinct from tarnishing (where red pigment is not fully suppressed by the inhibitor (silver) gene). In Bengals, excepting silvers and melanistics, a high degree of rufousing is desirable.
GLITTER, SATIN, GRIZZLE, TWEED, KARPATI AND MERLE
The Bengal Glitter Gene causes the golden glittered effect on Bengal cats and was inherited from a domestic cat used in the foundation stock. Glitter hairs have larger air cells in the core of the hair shaft. This oftens hair texture and produces an iridescent sheen to the coat. Glitter is inherited as an autosomal recessive. It is a mutation in gene Fgfr2. Breeders use the symbol "gl". Glitter type is related to colour: charcoal Bengals have silver glitter; "normal" tawny Bengals have gold glitter and snow/silver series Bengals have crystal glitter.
|
gl - Bengal Glitter Locus (posited) |
||
|
Genotype |
Description |
Phenotype |
|
Gl/Gl |
Homozygous - unglittered |
Normal coat. |
|
Gl/gl |
Heterozygous - unglittered carrying glitter |
Normal coat. |
|
gl/gl |
Homozygous - glittered |
Coat has sparkling glitter effect. |
There were thought to be 2 different types of Bengal glitter, referred to as "Mica glitter" (mi) and "Satin glitter" (sa) and both recessive. Mica glitter was believed to produce reflective hair-tips that looked as though they had mica (a highly reflective silicate crystal) embedded in the tip. Satin glitter was believed to affect the whole hair shaft. Researchers found only one glitter mutation in Bengals.
The Satin mutation is found in Tennessee Rexes. It affects the whole of the hair shaft and is found in many rodents (in some species, homozygous satin results in balding). The hair shaft is coated along its length with reflective material. This refracts light and give the coat a pearlised effect also termed "oyster". The hair is also soft, smooth and silky. In Tennessee Rexes it appears linked to the curled hair causing the "radical satin" effect. Straight haired variants have a less pronounced pearly appearance and appear to have some air pockets in the hairs. If satin and rexing are caused by one gene this pattern matches a recessive gene. If satin is a separate gene that could be inherited separately then the inheritance pattern also matches "dominant with additive effect" pattern.
|
Sa - Satin Locus (posited) |
||
|
Genotype |
Description |
Phenotype |
|
Sa/Sa |
Homozygous - unglittered |
Normal coat. |
|
Sa/sa |
Heterozygous - unglittered carrying satin |
Normal coat. |
|
sa/sa |
Homozygous - mica glittered |
Coat has satin/pearlised sheen. |
The Grizzle gene has been mainly seen in silver-tipped black Chausie has not been fully investigated. It appears to have been introduced from melanistic Jungle Cats (F chaus) in the same way that Glitter was introduced into the Bengal for its wild parents. Each grizzled hair has a black base with 5 bands of color ending in a black tip. Grizzle/silver tipped black is also seen in Jungle Cats and is a dominant gene and probably a mutation of either the agouti or extension gene. Although most noticeable on a black background, grizzle can occur with all the Chausie colourations.
Tweed is a recently noted colour mutation similar to roan in horses. White hairs are evenly intermixed with the solid base colour, such as black, to give a salt and pepper effect. Since all cats noted with this mutation were neutered rescue cats, it has not yet been investigated.
Karpati appears to be temperature sensitive, starting white with perhaps a darker strip down the spine, and then pigmented hairs (in any colour) start coming in making it appear gradually more and more coloured on the body. There are still unpigmented hairs interspersed amongst the pigmented ones, in a sort of roan effect. The extremities have very few of these pigmented hairs so look almost white in some, but paler at least in all, so pale mask, tails, paws, ears. (The name refers to the Carpathian region where this pattern is common.)
Merle occurs in red, cream and apricot silvers and results from uneven tipping. Visually it is similar to merle in dogs and mice and gives a solid base color with lighter patches resulting in a mottled or uneven speckled effect. Merle is better known in dogs where it also modifies the colour of the eyes, nose and paw pads. In cats it does not appear to be a merle mutation - solid/self reds have a residual tabby pattern because the non-agouti gene does not affect red pigment in the same way it affects black pigment. This can result in uneven tipping on cats with the silver inhibitor gene. Where the patches are large and irregular it is termed merle. Smaller more regular darker patches are also known as dapple.
COLOUR INHIBITOR GENES
The main colour inhibitor gene gives the cat a silver undercoat. The degree of colour inhibition creating the differences between silver tabbies, shaded and tipped silvers are believed due to the presence or absence of a separately inherited Wide Band gene that determines the width of silver/gold undercoat band caused by inhibitor gene. Cats with both the Inhibitor gene and the Wide band gene have a silver background colour while those with Wide band but no Inhibitor gene have a goldens background colour. The Inhibitor gene is more effective at blocking black-based pigment than blocking red-based pigment, hence there may be a breakthrough of reddish colour known as "tarnishing".
Dominant allele (I) produces that are fully colored only at the tip and have a white base. other genes influence the degree of tipping (the proprotion of each hair that is pigmented). Recessive allele (ii, also called i+i+) or no Inhbitor gene, combines with the agouti gene, to produce a golden undercoat. Poor (incomplete) expression of these or conflict with other genes results in tarnishing or excessively grey undercoat. Poor expression of non-agouti or over-expression of melanin inhibitor causes "smoke" cats to be excessively pale.
Another colour inhibitor that mimics silver has also been noted. The effect of wide band is variable suggesting there are multiple genes with a cumulative effect.
Dominant inhibitor + non-agouti = smoke series.
Dominant inhibitor + agouti + wide band = silver tabby/shaded silver/tipped silver series (variable expression).
Recessive inhibitor/no inhibitor + non-agouti = solid colour (there is no golden smoke series).
Recessive inhibitor/no inhibitor + agouti + wide band = golden tabby/shaded golden/tipped golden series (variable expression).
A "late colour change" silver-to-golden effect appears to involve the "sunshine" colour caused by the CORIN (Variable Wideband) gene. Kittens born as silvers may turn golden at 2-3 years old. Additionally, golden-from-birth cats may produce silver kittens.
|
I - Colour Inhibitor Locus |
||
|
Genotype |
Description |
Phenotype |
|
I/I |
Homozygous - Inhibitor - Silver |
Silver smoke/silver tabby/tipped (chinchilla)/shaded silver |
|
I/i (I/i+) |
Heterozygous - Inhibitor - carrier of non-inhibitor |
Silver smoke/silver tabby/tipped (chinchilla)/shaded silver |
|
i/i (i+/i+) |
Homozygous - Recessive inhibitor (non-silver) |
Non-silver (if wide band is present this gives Golden tabby/golden tipped/shaded golden) |
Wideband is a recessive trait with different Corin alleles involved in different breeds.
|
wb - Wide Band Locus (Corin Wideband gene is also called VWB - Variable Wideband) |
||
|
Genotype |
Description |
Phenotype |
|
wb/wb |
Homozygous - Wide Band |
Tipped |
|
-/wb or -/- |
Heterogygous or absent - Non-Wide-Band |
Tabby |
Wideband "Sunshine" is only seen in agouti (tabby patterned) Siberian cats (and possibly also Kurilian cats) where it produces reddish areas in the black, blue etc coat. Because Sunshine is on a different locus to the Inhibitor (silver) gene, it is possible for both genes to occur in the same cat resulting in a "bi-metal" appearance where both silver and golden areas are present.
|
wbSIB or vwbSIBor sg (Siberian Golden Colour) |
||
|
Genotype |
Description |
Phenotype |
|
- |
Homozygous for Non Sunshine - Normal colour |
Normal colour |
|
-/wbSIB |
Heterozygous - normal colour, carries sunshine |
Normal colour |
|
wbSIB/wbSIB |
Homozygous - sunshine colour |
Sunshine in non-silver cats, bi-metal in sunshine silver cats |
Wideband "Flaxen Gold" or "British Sunshine" is a very pale golden colour seen in British Shorthairs/Longhairs and Scottish Folds/Straights.
|
wbBSH or vwbBSH (British Shorthair/Longhair Flaxen Colour) |
||
|
Genotype |
Description |
Phenotype |
|
- |
Homozygous for Non Flaxen - Normal colour |
Normal colour |
|
-/wbBSH |
Heterozygous - normal colour, carries Flaxen |
Normal colour |
|
wbBSH/wbBSH |
Homozygous - Flaxen colour |
Flaxen in non-silver cats, extreme chinchilla in Flaxen silver cats |
Bengal sorrel is also a Corin wideband gene and follows the same inheritance pattern, just substitute wbBEN for wbBSH in the tables above. 12 Corin variants causing wideband effect have been identified in the Bengal.
HAIR LENGTH AND TYPE
In general, shorthair is dominant to longhair. Shorthaired cats sometimes produce longhairs due to a hidden recessive gene. Such carriers, and their offspring, are often removed from the breeding programme to eliminate undesirable recessives. Longhaired cats have 2 recessive genes for the longhaired trait and should always breed true for longhair. However, a rare autosomal shorthair gene that is recessive to longhair has been observed in some Persians lines where longhaired cats produced shorthaired kittens.
There appears to be a dominant gene in the York Chocolate breed determining absence or presence of undercoat. This gene may be present in other sleek-coated breeds (absence of undercoat would be an advantage in a hot climate).
The variability in longhairs is noted in the following:
|
L - Hair Length Locus |
||
|
Genotype |
Description |
Phenotype |
|
L/L |
Homozygous - Shorthair |
Shorthair (variable expression from Oriental type to Exotic shorthair type) |
|
L/l |
Heterozygous - Shorthair - carrying long hair |
Shorthair (variable expression from Oriental type to Exotic shorthair type) |
|
l/l |
Homozygous - longhair |
Longhair (variable expression from semi-long to Persian type) |
|
ls - Recessive Shorthair Locus (provisional gene symbol) |
||
|
Genotype |
Description |
Phenotype |
|
Ls/Ls |
Homozygous - Longhair |
Longhair |
|
Ls/ls |
Heterozygous - Longhair - carrying short hair |
Longhair |
|
ls/ls |
Homozygous - Shorthair |
Shorthair |
|
Yuc - York Chocolate Undercoat Locus (posited) |
||
|
Genotype |
Description |
Phenotype |
|
Yuc/Yuc |
Homozygous - lacks undercoat |
Sleek fur, no undercoat |
|
Yuc/yuc |
Heterozygous - lacks undercoat |
Sleek fur, no undercoat |
|
yuc/yuc |
Homozygous - full undercoat |
Undercoat present |
HAIRLESS AND REX MUTATIONS
In addition to the genes listed here, there are a number of hairless mutations and rex mutations, both extinct and extant, whose mode of inheritance is not known. Many of these unknown genes may be identical to currently bred mutations, while others will be novel mutations. The effect of rex and wirehair genes also depends on the coat length and probably on polygenes. Where two rex genes on different loci are present, these may interact to produce a coat type different to either parent.
The Canadian hairless gene is recessive to normal hair, but is dominant to Devon Rex (in 2010 DNA analysis found that Devon Rex Sphynx genes are alleles of the same gene, previously they were believed to be different loci with Sphynx being epistatic to Devon Rex). The order of dominance at the Hairless-Devon locus is Hr > hr > re. (The gene involved is KRT71 and the order of dominance is KRT71+ > KRT71hr > KRT71re - "+" indicates "wild type")
Two other extinct recessive hairless genes have been noted, but the locus not identified: French Hairless (h) and Redcar (UK) Hairless (hd). The Russian Hairless gene is an incomplete dominant hairless trait found in the Peterbald and Don Sphynx where an initial rex coat may be present and hairlessness develops progressively during the first 24 months; test matings indicate th presence of 2 modifers.
The extremely rare Kohana Cat (Hawaiian hairless) was believed to be due to the presence and interaction of both the dominant Russian Hairless allele and the recessive Canadian Sphynx allele or alternatively to a new dominant gene mutation. DNA analysis in 2010 found that the Hawaiian hairless was the same mutation as Canadian Sphynx. The analysis also found that Sphynx and Devon Rex are different mutations of the same gene. The relationship between these mutations from most dominant to most recessive is: Russian Hairless >> Normal Hair >> Sphynx >> Devon Rex
Wirehair is a dominant allele with incomplete penetrance i.e. cats with the dominant wirehair allele (genetically wirehair cats) may have a normal coat, but produce wirehaired offspring.
|
r - Cornish/German Rex Locus |
||
|
Genotype |
Description |
Phenotype |
|
R/R |
Homozygous - Straight haired cat |
Standard coat - no effect |
|
R/r |
Heterozygous - Straight haired cat carrying - C rexing |
Standard coat - no effect |
|
r/r |
Homozygous - C rexed cat |
Rexed - wavy fur |
|
re - Devon Rex Locus |
||
|
Genotype |
Description |
Phenotype |
|
Re/Re |
Homozygous - Straight haired cat |
Standard coat - no effect |
|
Re/re |
Heterozygous - Straight haired cat carrying - D rexing |
Standard coat - no effect |
|
re/re |
Homozygous - D rexed cat |
Rexed - wavy fur |
|
ro - Oregon Rex Locus |
||
|
Genotype |
Description |
Phenotype |
|
Ro/Ro |
Homozygous - Straight haired cat |
Standard coat - no effect |
|
Ro/ro |
Heterozygous - Straight haired cat carrying - O rexing |
Standard coat - no effect |
|
ro/ro |
Homozygous - O rexed cat |
Rexed - wavy fur |
|
Se - Selkirk Rex Locus |
||
|
Genotype |
Description |
Phenotype |
|
Se/Se |
Homozygous - S Rex cat |
Rexed - wavy fur, finer, curlier and sparser than heterozygote |
|
Se/se |
Heterozygous - S Rexed cat - carrying normal coat |
Rexed - wavy fur, preferred coat type |
|
se/se |
Homozygous - Straight haired cat |
Standard coat - no effect |
|
Lp - Laperm Locus |
||
|
Genotype |
Description |
Phenotype |
|
LP/LP |
Homozygous - LP Rex cat |
Rexed - wavy or ringletted fur |
|
LP/lp |
Heterozygous - LP Rexed cat - carrying normal coat |
Rexed - wavy or ringletted fur |
|
lp/lp |
Homozygous - Straight haired cat |
Standard coat - no effect |
|
Wh - Wirehair Locus |
||
|
Genotype |
Description |
Phenotype |
|
Wh/Wh |
Homozygous - Wirehair |
Bristly, wiry fur |
|
Wh/wh |
Heterozygous - Wirehair carrying - staight hair |
Bristly, wiry fur |
|
wh/wh |
Homozygous - Straight hair |
Standard coat - no effect |
|
hr - Canadian Hairless (Sphynx) Locus |
||
|
Genotype |
Description |
Phenotype |
|
Hr/Hr |
Homozygous - Coated cat |
Standard coat |
|
Hr/hr |
Heterozygous - Coated cat - carrying Sphynx hairless allele |
Standard coat |
|
hr/hr |
Homozygous - Sphynx Hairless cat |
hairless cat |
|
Hrbd- Russian Hairless Locus + Theoretical MInimiser and MAximiser Modifiers |
||
|
Genotype |
Description |
Phenotype |
|
Hrbd/Hrbd |
Homozygous - Hairless |
Hairless cat |
|
Hrbd/Hr |
Heterozygous - Hairless -carrying normal coat |
Hairless cat |
|
Hr/Hr |
Homozygous - Normal Coated cat |
Standard coat - no effect |
|
Donskoy coat genetics is poorly understood. Below are theoretical modifiers affecting expression of Hrbd. These were based on observation of one breeding line only and other breeders have seen different results |
||
|
HrHr + hiha |
normal haired (assume normal-haired cats have one of each modifier) |
|
|
HrbdHr + hihi |
Brush-coat |
|
|
HrHr + hiha |
normal haired (assume normal-haired cats have one of each modifier) |
|
|
HrbdHr + hiha |
brush coat with some bald areas |
|
|
HrbdHr + haha |
brush coat that turns bald |
|
|
HrbdHrbd + hihi |
velour coat |
|
|
HrbdHrbd + hiha |
flocked coat |
|
|
HrbdHrbd + haha |
rubber bald born bald and remains bald |
|
|
If these modifiers were correct, (HrbdHrbd + haha) x (HrbdHrbd + haha) (both parents rubber bald, born bald) should only produce bald kittens that remain hairless. Matings of 2 rubber bald, born bald parents have produced kittens with other hair types. |
||
EARS
The curled ear trait in American Curls and derivative breeds is a dominant trait with variable expression. A curled ear mutation was observed in a female stray Australian cat. She did not pass this on to her offspring. Back-crossing to identify a possible recessive curled-ear trait was impossible as the mother became ill and had to be spayed.
Forward folded ears is a dominant trait with incomplete penetrance and linked to disabling skeletal problems, especially in homozygotes. Incomplete penetrance means a prick-eared cat may genetically be a fold-eared cat.
|
Cu - Curled Ears Locus |
||
|
Genotype |
Description |
Phenotype |
|
Cu/Cu |
Homozygous - Curled ears |
Ears Curl backwards (variable expression) |
|
Cu/cu |
Heterozygous - Curled Ears - carrying normal ears |
Ears Curl backwards (variable expression) |
|
cu/cu |
Homozygous - Normal ears |
Standard ears - no effect |
|
Cua - Australian Curl Ears Locus (posited) |
||
|
Genotype |
Description |
Phenotype |
|
Cua/Cua |
Homozygous - Prick ears |
Normal upright ears |
|
Cua/cua |
Heterozygous - Prick Ears - carrying curled ears |
Normal upright ears |
|
cua/cua |
Homozygous - Curled ears |
Ears Curl backwards |
|
Fd - Folded Ears Locus |
||
|
Genotype |
Description |
Phenotype |
|
Fd/Fd |
Homozygous - Folded Ears |
Ears Folded Forwards |
|
Fd/fd |
Heterozygous - Folded Ears - carrying non-folded ears or standard ears (rare) |
Ears Folded Forwards |
|
fd/fd |
Homozygous - Prick-ears |
Prick-ears |
EYES
Many genes govern eye colour, creating shades ranging from blue to green to yellow to orange and many shades inbetween. The epistatic white gene can cause blue eyes as can the white spotting gene where there is high grade white spotting. The Siamese colourpoint allele produces blue eyes while the Burmese Sepia allele has orange eyes; cat with one of each (Tonkinese) having aqua eyes. Blue-eyed and pink-eyed albinos also occur and are part of the colourpoint series of alleles.
A notable gene affecting eye colour is that in the Ojos Azules. This is a dominant mutation giving blue eyes, regardless of the coat colour, and white splashed extremities. Other blue-eyed mutations have been observed, but not selectively bred.
|
Oj - Ojos Azules Locus (no official gene symbol assigned?) |
||
|
Genotype |
Description |
Phenotype |
|
Oj/Oj |
Homozygous - Ojos Azules eye colour |
Blue eyes, white splashed extremities. |
|
Oj/oj |
Heterozygous - Ojos Azules carrying other eye colour |
Blue eyes, white splashed extremities. |
|
oj/oj |
Homozygous - non Ojos Azules eye colour |
Any other eye colour, any markings on extremities are due to other White Spotting genes |
TAILS
The normal tail is full-tailed. There are 2 identified tail mutations. Japanese Bobtail, found throughout Asia is a recessive. The Kurilian s bobbed tail is due to a dominant gene with incomplete penetrance, while the similarly named Karelian s bobbed tail is a recessive gene. These are not associated with any skeletal abnormalities. Manx taillessness is a dominant trait with variable expression ranging from from tailless (Rumpy) through rumpy riser, stumpy and longy. It is a classic lethal gene and is associated with skeletal and spinal problems (these may be due to "linked genes" as breeders have reduced the incidence of Manx syndrome).
|
Jb - Japanese Bobtail Locus |
||
|
Genotype |
Description |
Phenotype |
|
Jb/Jb |
Homozygous - Normal tail |
Normal tail |
|
Jb/jb |
Heterozygous - Normal tail carrying bobtail |
Normal tail |
|
jb/jb |
Homozygous- Japanese Bobtail |
Short curled bobtail |
|
Kur - Kurilian Bobtail Locus |
||
|
Genotype |
Description |
Phenotype |
|
Kur/Kur |
Homozygous - Bobtail |
Short curled bobtail |
|
Kur/kur |
Heterozygous - Bobtail tail carrying normal tail |
Short tail, may or may not be curled |
|
kur/kur |
Homozygous- Non Kurilian Bobtail |
Normal tail |
|
M - Manx taillessness Locus |
||
|
Genotype |
Description |
Phenotype |
|
M/M |
Homozygous - Tailless, die in utero |
Die in utero, gross neural tube defects. |
|
M/m |
Heterozygous - Tailless - carrying normal tail |
Ranges from no tail (rumpy) to short tail |
|
m/m |
Homozygous - Normal tail |
Full-length tail |
LIMBS AND PAWS
The Munchkin mutation affects limb length only. It is not achondroplastic dwarfism as the cranium is unaffected. It is probably hypochondroplasia or pseudochondroplasia. Munchkin is an autosomal dominant, possibly lethal in homozygous state as breeders have reported litters smaller than average.
Classic (mitten-foot/thumb-cat) polydactyly is dominant in action with variable expression and incomplete penetrance. This gene has only cosmetic effect ranging from one or more extra toes, usually in mitten-foot conformation. Due to incomplete dominance, some normal footed cats are genetically polydactyl. Other polydactyl genes with other modes of inheritance are probable; so far the Radial Hypoplasia gene has been identified as one cause of patty-foot polydactyly. Radial hypoplasia ranges from cosmetic (polydactyly) to debilitating (absent radius) and patty-foot cats are not selectively bred. Other polydactyl mutations, both dominant and recessive, are likely.
The identified form of Syndactyly (split foot) in cats is a dominant gene resulting in a two-digit paw that resembles a lobster-claw. It is considered debilitating and is not selectively bred. There are probably phenotypically similar conditions, due to other mutations, which have not been investigated.
|
Mk - Munchkin |
||
|
Genotype |
Description |
Phenotype |
|
Mk/Mk |
Homozygous - Short legs |
Munchkin |
|
Mk/mk |
Heterozygous - Short legs |
Munchkin |
|
Mk/mk |
Homozygous - Normal legs |
Normal length legs |
|
Pd - Polydactyl Locus |
||
|
Genotype |
Description |
Phenotype |
|
Pd/Pd |
Homozygous - Polydactyl |
Extra toes |
|
Pd/pd |
Heterozygous - Polydactyl - carrying normal number of toes |
Extra toes |
|
pd/pd |
Homozygous - Normal number of toes |
Standard number of toes - no effect |
|
Sh - Split Foot Locus |
||
|
Genotype |
Description |
Phenotype |
|
Sh/Sh |
Homozygous - Syndactyl |
Split foot (lobster claw syndrome) |
|
Sh/sh |
Heterozygous - Syndactyl carrying normal number of toes |
Split foot (lobster claw syndrome) |
|
sh/sh |
Homozygous - Normal number of toes |
Standard number of toes - no effect |