
THE DOMESTICATION OF THE CAT
Cats can live as cherished domestic pets or as tolerated wild/half-wild animals living in proximity to human dwellings. Domestic pets generally rely on their human benefactors for food and shelter, while the wild/half-wild cats live as self-sufficient hunters or scavengers. Domestic cats that revert to the wild state are known as feral cats (semi-feral in the case of half-wild cats).
Biologists consider cats to be "exploitive captives" - captives of humans, but exploiting this relationship to their own ends rather than suffering from it. Throughout its history, cats have also been symbiotes where both humans and cats gain from the relationship - humans gain rat-catchers while cats gain shelter and supplementary feeding. Elsewhere, where rodent control is not an issue, they are commensals - present alongside man and dining on man's leftovers or on garbage (on food scraps that would otherwise attract vermin or disease). Because of these roles, cats and humans have a long history together. The earliest archaeological evidence of this relationship dates comes from 10,000 years ago (clay figurines in Syria, Turkey and Israel) and 9,500 years ago (evidence of cats living alongside humans in Cyprus).
Rather than viewing humans as either predator or potential prey, at some point in prehistory, a small wildcat species found itself a niche in human communities by living on the rodents attracted by agricultural activities. In doing so, it performed a beneficial service to humans and became a welcomed domestic animal rather than a tolerated wild animal.
Ancestral Species
The ancestral species of the domestic cat has been the source of much debate among naturalists and cat fanciers. H C Brooke wrote a contribution to Frances Simpson's "The Book of the Cat" (1903, "Some Foreign Cats") mentioning the Egyptian Cat (Felis maniculata) and the European Wild cat (F catus) (the Latin names in use at that time):
"It might reasonably be imagined that our common cat was derived from the last-named, considering that at one time it was a common animal all over England as well as on the continent. The untameable ferocity of this variety - which is probably the least amenable of all living creatures - has doubtless prevented its ever having been domesticated and the high value which, as we learn from old writings, was a common animal in England, plainly show that F catus was not the ancestor of F domestica, although the two will freely interbreed. Many years ago, the old Spanish wild cat which used to be kept at the Zoological Gardens in the so-called aviaries, now occupied by the civets, mated with his cage mate - a tortoiseshell-and-white queen - and of these cross-bred kittens both Sir Claud Alexander and the writer of these lines [Brooke] possessed specimens. It is usually assumed that the Egyptian or Caffre cat is the progenitor of the majority of the domestic cats [ ] it is found over the whole of Africa, and it is quite easy to understand how, with its eminently tameable disposition, it gradually spread over Europe. Our so-called Abyssinian cats [ ] bear a very striking resemblance to this handsome variety of cat."
In 1907, the taxonomist Pocock described two types of domestic cat F torquata (striped tabby) and F catus (blotched tabby) although F catus was elsewhere used to describe the European wildcat. Linnaeus assigned the name F catus to the domestic blotched tabby. The domestic cat has also been known as F domestica.
Pocock (1907) noted the accepted ancestor of the domestic cat is the Arabian and African form of the wildcat, Felis lybica (F lybica). The European wildcat (including the Scottish wildcat) is F silvestris. These two wildcats interbreed easily and are currently considered related subspecies properly called F silvestris silvestris (a robust form adapted to colder climes) and F silvestris lybica (J Clutton Brock, A Natural History of Domesticated Mammals, 106-112, 1987). The prehistory of domestic cats is complicated and it is suggested that species other than F s lybica were also involved e.g F s silvestris (European wildcat) and F ornata (Asiatic wildcat).
F lybica has a very large geographic range and several subspecies of variable appearance. These subspecies can interbreed to produce offspring whose appearance is intermediate between the parents. Generally speaking, moving from north to south there is a gradation of coat thickness, intensity of ground colour and the amount of "tabby" markings (Robinson 1991), with northern subspecies being more heavily built, thicker furred with a rounded-tipped tail and darker, more distinct tabby markings than southern subspecies. The African wildcat has a lighter build, less distinct markings and thin, tapering tails and varies from rusty, through sandy and tawny to grey with faint tabby markings.
In 1951, Pocock described 26 subspecies of wildcat though many were probably variants or intermediates since only 4 groups are recognised today. Weigel (1961) and Hemmer (1978) described four groups: the "forest" cats (silvestris) of Europe, the Caucasus and Asia Minor; the "steppe" cats (ornate) of South and Central Asia; the "tawny" cats (lybica) of Africa and the Middle East; and the familiar domestic cat (catus).
As late as 1996, and in spite of the hybridisation at London zoo reported by H C Brooke (1903), researchers have claimed that F catus was not closely enough related to F s silvestris to produce fertile hybrids. In fact they hybridize naturally in Scotland and the local sub-species of F silvestris, F s grampia (Scottish Wildcat) is in danger of being supplanted by hybrids and mongrels.
The Role of Other Small Species in Domestic Cat Evolution
While generally accepting F s lybica as the main ancestor, there was continued debate into the role of other small cat species.
A species credited with partial ancestry of the domestic cat (especially of those in India) was F chaus (jungle cat, marsh cat, reed cat) which was present in Ancient Egypt and possibly depicted in Ancient Egyptian art. There is some evidence that F chaus was kept alongside F s lybica in Egyptian temples during the time that cats were traditionally believed to have been domesticated. The two species interbreed successfully, but there is no compelling evidence for past hybridization between the two species. The Egyptians kept many wild animals so the presence of of F chaus remains should not be considered proof of hybridisation. In addition, recent genetic evidence indicates domestication and spread of the domestic cat began earlier than previously thought.
F chaus produces fertile offspring with domestic cats, but its contribution to the ancestral gene pool, if at all, is minimal. Tabor believed its influence was greater than admitted and has largely been ignored (R Tabor, Cats - The Rise of the Cat, p16, 1991), but this is not upheld by modern genetic studies. Tabor stated F catus x F chaus hybrids are far more docile than F catus x F s silvestris hybrids; suggesting that F chaus contributed genes that made early domestic cats humanisable. However, F catus x F s lybica hybrids are also more docile than F catus x F s silvestris hybrids so "docilising genes" could originate from F s lybica. Hybrid offspring of domestic cats (or African Wild cats) and European Wild cats take after the European Wild cat (i.e. non-docile) and will move away from man. The heavy body type of British cats and Persians was once thought to be due to the genetic influence of the European wildcat.
Tabor suggested other species may have contributed to the mix of genes in the domestic cat, accounting for some traits not found in F s lybica, F s silvestris or F chaus. These species include the golden cat (F temminckii) which occurs between south China and Sumatra and which, he suggested, may have contributed towards the oriental type cats. The link between the Siamese and Temminck's cat appears due to naming confusion in the early 1900s. In Ceylon, the Siamese cat was known as "Gould's Cat", having been introduced there by a Mr Gould. The Burmese Sacred Cat (the Burmese, not the Birman) was known to early British cat fanciers as the "Gold Cat". A wild cat of the region was known as the "Golden Cat" (Temminck's Golden Cat) or "Bay Cat". HC Brooke (1927), believed these similarities of name to be the reason that Temminck's Golden Cat (or Bay Cat) was claimed to be an ancestor of the Siamese.
18th Century German naturalist Peter Pallas recorded that Pallas's Cat (F manul) found in Iran through to western China would breed with domestic cats and might have contributed the longhair trait. It is sometimes suggested that Persian longhairs may be partly descended from Pallas's cat, although there is no skeletal evidence for this. Similar suggestions have been made regarding the Sand Cat (F margarita) as an ancestor of longhairs. This was discounted even before DNA studies because the domestic cat does not have long fur on the underside of its paws.
Others suggestions include the leopard cat (F bengalensis) from eastern Siberia through to South-east Asia, and the sand cat (F margarita) found from the Sahara to Baluchistan. In addition, the naturally occurring Russian "Ussuri" breed is believed to be derived from Asian Leopard Cat/domestic cats, but this is based on appearance only.
Detailed studies of skulls, skeletons, anatomical features and, more recently, genetics do not support theories that the cat is a complex hybrid of multiple species. Domestic cats are anatomically and skeletally similar to F s lybica. Chromosome counts and modern hybrids show that it is feasible for Pallas's cat, the Leopard cat and the Asian Golden cat to have contributed in the past, but the domestic cat is most similar in type to the African wildcat and variations in appearance (colour, fur length) are due to genetic mutations.
In 2007 studies published in Science journal of mitochondrial DNA from cats in the UK, US, Germany, Israel, Spain and France showed that at least 5 female ancestors from the Egyptian region gave rise to all the domestic cats alive today. Apart from accidental cross-breeding, European wildcats have not contributed to the domestic cat's ancestry. It also ruled out the Central Asian wildcat, the Southern African wildcat and the Chinese desert cat as ancestors.
While Brooke correctly identified the Egyptian or Caffre (now known as F s lybica) cat as progenitor of "the majority of the domestic cats", he incorrectly believed that other countries' domestic cats were descended from their indigenous smaller wild cats. According to the naturalist Blyth, quoted by Brooke, this resulted in the two varieties of Indian domestic cats: the fulvous (ticked) variety derived from the Jungle cat (F chaus) and the spotted varieties supposedly derived from the Leopard cat, Desert cat and Rusty Spotted cat. To illustrate this belief in different races of domestic cats in India, Brooke wrote of an Indian cat acquired by his wife from a London shoemaker who had acquired from a sailor who had stolen it from a hotel in Bombay.
"Very curious and handsome is the Indian cat[ ] His most striking peculiarities are the length and slenderness of his limbs, the extreme shortness of his coat, and his thin and tapering tail [ ] His ears are small, but as a kitten they were of enormous size, and with his long and pointed head gave him a most weird experience. The voice of this cat is very variable, and far more resembles the raucous call of the Siamese than the voice of any European cat." In retrospect, this is an Oriental cat rather than a variety descended from indigenous wildcats, but in Brooke's time only the only recognised Oriental was the Siamese. The variety had probably reached Bombay the same way it left - on shipboard from Thailand or the Malay Peninsula. The Siamese of that time was far less extreme than the modern variety and the illustration shows the Indian cat as similar to an Abyssinian.
In the US, the cat may possibly hybridize with F rufus (Bobcat) and has been deliberately hybridized with F chaus, F bengalensis, F geoffroyii (Geoffroy's cat), F viverrina (Fishing Cat) and F serval (Serval) and Margay (possibly also with the ocelot). Future generations of some varieties of F catus will be more complex hybrids than their predecessors.
DNA Studies
Fossil evidence of cats is hard to interpret, but recent DNA studies indicate that the 37 species of modern cats have their roots in South East Asia 11 million years ago. Eight major lineages emerged, with the domestic cat being most closely related to the African and European wild cat and the Chinese desert cat according to a study of mitochondrial (maternal) DNA and X and Y chromosomal DNA by the National Cancer Institute in Frederick, Maryland, USA. Scientists identified distinctive genetic clades (groups). One clade includes domestic cats and some wildcats from the Middle East, indicating a close relationship between these species.
Of the other lineages, the Panthera lineage ("roaring cats" such as the lion, jaguar and tiger, emerged first). Next came a group of three Asian species (Bay cat, Asian golden cat and Marbled cat). Then three African Species (Caracal, African Golden cat and Serval) followed by the New World Ocelot. The remaining 4 lineages branched more recently: the Lynx, Puma, Leopard cat and domestic cat. The common ancestor of modern cats migrated from Asia into Africa 6-8 million years ago (Caracal lineage). Cats crossed the Bering land bridge to North America about 8 million years ago with some migrating into South America by the Panamanian land bridge.
Of the various lineages, the African Wildcat (descended from F lumensis Martelli's Wildcat about 600,000 to 900,000 years ago) would be the one to give rise to the familiar domestic cat and it may have been domesticated more than once although modern cats are traceable to a single domestication event.
An analysis of the spread of domestic cats and the relationship between pedigreed breeds was published in October 2007 (M.J. Lipinski, et al., The ascent of cat breeds: genetic evaluations of breeds and worldwide random-bred populations, Genomics (2007)). The researchers used genetic markers to trace cats from the presumed site of domestication to distant regions of the world and also to determine if breeds came from their purported place of origin. The sample comprised genetic data from 1100 cats across 5 continents, including 22 pedigree breeds and 17 random-bred populations (Brazil, Hawaii, New York, Texas, Germany, Finland, Italy , Turkey , Israel, Egypt, Tunisia, Kenya, Sri Lanka, Singapore, Vietnam, Henan (China), Korea).
The breeds used in the study were 15 of the 16 CFA foundation ("natural") breeds (Abyssinian, American Shorthair, Birman, British Shorthair, Burmese (i.e. American Burmese), Chartreux, Egyptian Mau, Korat, Maine Coon, Norwegian Forest, Persian (presumably excluding Colourpoint Persian), Russian Blue, Siamese, American Turkish Angora, American Turkish Van), 2 CFA breeds under development (Siberian and Sokoke) and 5 more recently developed CFA breeds (Exotic Shorthair, Havana Brown, Japanese Bobtail, Singapura and Sphynx).
DNA samples mostly obtained from cat shows and from cat owners in the USA. Additional Korat, Turkish Angora, Turkish Van, and Siberian samples were acquired from Europe where the breeds have the same standards. The study excluded first-degree relatives and minimised shared grandparentage (this cannot be eliminated in breeds with small gene pools). Wild/domestic hybrids (e.g. Bengal) were not included in the study. Turkish Angoras and Turkish Vans bred in their native country were excluded.
The random-bred cats included feral cats living independently of humans, random-bred cats that had human assistance (e.g. semi-ferals) and random-bred pet cats. Littermates and parent/kitten relationships were excluded from the study. The wild species used in the studies were: African wildcat (F. s. caffra), European wildcat (F. s. silvestris), Arabian wildcats (F. s. tristami from Israel).
Note: The researchers failed to acknowledge that the American showbench versions of the Turkish Angora and Turkish Van had been created by crossing Persians to other cats. While the American Angora broadly resembles the ancestral Turkish breed, it is genetically mongrelised and not representative of cats in Turkey.
Why The African Wildcat?
Why was the African wildcat domesticable while its more northern cousins were (and are) not? As well as colouration among subspecies across its range, temperament also varies. The more northerly F silvestris is generally intractable even if hand-reared. F lybica is generally more amenable to human presence. This would be a factor during the early domestication of the cat.
African wildcats hunt a variety of prey animals, but rarely scavenge carrion. Unlike feral domestic cats, which sometimes form colonies, African wildcats are solitary. Domestic cats will often eat carrion and will form colonies where there is a rich food supply e.g. rubbish dumps, farmyards, but prefer to remain solitary where prey is more thinly distributed (Liberg and Sandell 1988). Studies in Saudi Arabia indicate that wildcats continue to remain solitary while feral cats formed colonies around a garbage dump.
However, African wildcats do have some ability to co-operate since captive female wildcats have been seen to assist mothers in supply food for kittens (Smithers 1983), a behaviour observed in feral cat colonies. Early domestication is believed to be due to humans attracting rodents which attracted cats. Those wildcats which best tolerated the presence of other cats would have been better able to exploit the food supply and would have produced young which inherited their more sociable tendencies. In addition to becoming more sociable, cats also evolved to exploit humans by scavenging carrion and food scraps.
Neolithic Remains (Origin of Domestic of Cats in the Levantine Region)
"Levantine" refers to Cyprus and land bordering the Eastern Mediterranean Sea in West Asia, including Syria, Lebanon, Jordan, Palestine, Israel, and south Cilicia (Turkey). The "Fertile Crescent" includes the Levant, Ancient Mesopotamia, and Ancient Egypt. The Levant is the land bridge between Africa and Eurasia.
Remains of cats have been found from the Pre-pottery Neolithic of Jericho (circa 7000 BC) though these may have been hunted animals or the equivalent of modern feral cats, attracted to food scraps and tolerated at best. Tamed cats have possibly lived in association with humans far earlier than archaeological and historical records imply. Cat remains (bones, teeth) are often retrieved from prehistoric sites, but it is impossible to know whether these were companions or prey (pelts, meat). Stone and clay figurines of cats have been found in Syria, Turkey and Israel. These 10,000 year old sculptures indicate that cats were important to those cultures in ways other than a source of food. The process of domestication may have begun 10,000 years ago rather than only 4000 years ago.
Initially, cats were probably tolerated by humans because they killed the mice and rats that were attracted to human food stores. The cats themselves may have been eaten or hunted for their pelts. Later, humans would have deliberately encouraged the presence cats as rat-catchers instead of hunting them.
In Cyprus, African Wild cat remains (based on mitochondrial DNA) dated to around 5000 BC have been found and more recent finds are even older - 9,500 years. This is interesting because an African wildcat would have to have been transported to Cyprus by humans. Cyprus has never had any indigenous wildcats. The bones of cats, mice, and humans were found together and all three of these species first appear on Cyprus at the same time. This suggests that humans brought mice to the island of Cyprus by accident and the cats on purpose to control the mice, suggesting that the cat's value as a rodent-catcher was already recognised. Analysis of nucleus DNA found that the cats were hybrids of European and African wildcats, and the mitochondrial DNA came from a maternal African wildcat ancestor.
Studies, based on the discovery of a complete cat skeleton buried alongside a Neolithic human at Shillourokambos, Cyprus, suggested that domestication of cats began 9,500 years ago. The cat and human skeletons were positioned symmetrically with the heads pointing west (towards the setting sun) and were in an identical state of preservation i.e. buried at the same time. The cat in the grave may have been "pre-domesticated", a halfway stage between being wild and being domesticated, or possibly a pet. The person was about 30 years old at death and other grave goods, such as stone tools and seashells, suggest high status; s/he may have had a special relationship with cats: it may have had religious significance, been a totem animal or an exotic pet.
During the Neolithic period, agriculture was beginning to spread from the Near East and grain storage would have attracted large mice populations. Cats may have been encouraged to settle in villages to control the mice. The cat resembled the African wildcat (F s lybica), a species whose kittens are amenable to taming if taken as kittens, so it is possible that an abandoned kitten, or a particularly attractive one, was adopted as a pet. This may be analogous to the keeping of exotic animals today (status symbol or novelty) rather than domestication of a whole species. DNA studies published in 2025 (M. De Martino et al, Science, 27 Nov 2025) challenged the theory of a widespread Neolithic introduction of domestic cats to Europe, instead placing their arrival in Europe several millennia later.
While the cat may have been a domestic or exotic pet, it was killed at 8 months old in order to be buried with its "owner". It was buried intact, with no signs of butchering, suggesting it was treated as an individual or companion, and not as a food offering. Burnt cat bones from a similar period at the site show that cats formed part of the Neolithic diet in the region, but the presence of cat remains does not indicate a widespread introduction of cats into Europe.
Origin of Domestic of Cats in the Middle East
The early movement of cats along trade routes makes it difficult to determine exactly when and where the cat was first domesticated. Genetic studies published in Science journal in 2007 indicate origins in the Middle East's Fertile Crescent (present-day Iraq, Syria, Lebanon and Israel), probably to early farming communities which attracted rodents. Archaeological studies suggested a single domestication event in the Near East approximately 9000–10,000 years ago. Scientists acknowledge that cats (perhaps in a semi-domesticated state) may have lived in close proximity to humans when humans formed permanent agricultural settlements around 10,000–11,000 years ago in the Fertile Crescent. Modern domestic cats would then be descended from cats domesticated in the Mediterranean region.
Early genetic studies show that the ancestors of the domestic cats broke away from their wild relatives and started living with humans 100,000 – 130,000 years ago, much earlier than previously thought. This much earlier date for domestication supposedly accounted for the early presence of domestic cats in Cyprus, Pakistan, North Africa and elsewhere; cats were already domesticated (or partially so) before they were revered in Ancient Egypt. Brian Vesey-Fitzgerald "Cats" (1958) wrote "I think that [domestication] probably happened in many different parts of the world. It may not even have happened first in Africa. There are mentions of the cat in Sanskrit writings of about three thousand years ago which suggest that it may have been domesticated in India at about the same time as in Egypt. And it was certainly domesticated in China long before the birth of Christ. However, it is from Egypt that we have our first definite knowledge of the cat as a domestic animal."
Ancient mitochondrial DNA (mtDNA) data supported the dual-origin model (Neolithic Levant around 9500 years ago and Pharaonic Egypt around 3500 years ago) for cat domestication and spread, indicating an initial spread from Anatolia to Europe by Neolithic farmers approximately 6400 years ago, followed by a second wave from Egypt around 2000 years ago. A later genetic study using nucleus DNA (M. De Martino et al, Science, 27 Nov 2025) challenged this. It found that the cat remains from the Neolithic and Copper Age (9500 to 6300 years ago) in southeast Europe and Anatolia, which had previously been identified as F. l. lybica (African wildcat) based on their mitochondrial, were actually F. silvestris wildcats whose ancestors had hybridized with F. l. lybica cats. Analysis of ancient cat remains showed that degree of African wildcat ancestry ranged from 9% in Bulgaria to 34% in central Anatolia.
The earliest F. l. lybica/F. catus cat in Europe was from Sardinia (Italy) about 2200 years ago. It was genetically similar to modern wildcats in Sardinia, and closely related to a F. l. lybica wildcat from Morocco. All other archaeological F. l. lybica/F. catus cats from Europe and Anatolia dated from approx. 2000 years ago onward. Their gene pool was typical of modern domestic cats and closer to modern African wildcats than to archaeological Levantine cats. The maternal DNA shows rapid their dispersal across the Mediterranean in the past 2000 years.
Even after the 2025 DNA study, it is uncertain whether domestic cats originated in the Levant, Egypt, or elsewhere in the natural range of African wildcats. They appear to have been introduced to Europe around 2000 years ago, probably from North Africa. M. De Martino et al (2025) point to 2 separate dispersals of cats from Africa into Europe. The first dispersal of was the earlier introduction (1000 BCE – 1 BCE) of wildcats from Northwest Africa may have been given rise to the present-day wild population in Sardinia, but not to modern domestic cats (African wildcat populations in the Mediterranean islands of Sardinia and Corsica do not appear to be feral domestic cats, but a separate wildcat lineage.) The second dispersal involved a distinct population from North Africa and led to modern domestic cats in Europe around 200 years ago.
Cats then spread between Old World civilizations along trade routes, but remained little changed from their wild ancestors (if compared to dogs). Different studies have suggested likely feline dispersal routes based on coat colour mutations (e.g. classic tabby, ginger) and structural mutations (e.g. polydactyly, bobtails).
Interbreeding between feral cats, tame domestic cats and some wild subspecies of Felis silvestris (and occasionally with other wild species) has not greatly impacted the cat's form or behaviour and they remained the major small carnivore living around human settlements (ferrets and mongooses have been used for similar roles, but to a limited degree). Feral cat populations were tolerated, or encouraged, for pest control with no need for selective breeding to specialise them for particular roles as happened with dogs, sheep and other domestic animals. When artificial selection took place (from the late 19th Century onwards in Western Europe, anecdotally earlier in Thailand), it was on the basis of aesthetics, not function.
Domestication of Cats in North Africa
Nineteenth century explorers of North Africa have recorded that local people caught and reared African Wild Cats as pets, keeping them around their huts and enclosures. Similar events have been reported (right up to modern times) in North African and Arabia.
The Azande tribe of southern Sudan live in tolerant proximity with the African Wild cat, though they haven't domesticated it. The cats are content to find shelter and scavenge food around the tribal settlements and show little fear of people. In this it is much like the urban fox in Britain - tolerates proximity with humans but not tame.
The relative docility of the African Wild cat and its ability to leave close to people without being held in captivity, suggest that the cat itself moved in with people exploiting shelter and food supplies in and around human settlements.
Domestication of Cats in Ancient Egypt
According to J A Baldwin, there was little question that European domesticated cats came out of Egypt between 3,000 BC and 2,000 BC (J A Baldwin, Anthropos 70:428-448, 1975). There are no confirmed records of domesticated cats from the Old Kingdom (2686-2181 BC). The earliest Ancient Egyptian evidence of the presence of cats dates from about 2500 BC and there was evidence of primitive domestication around 2000 BC, however the Egyptians also depicted many non-domestic species. There was little pictorial evidence to suggest that cats were fully domesticated until the New Kingdom, 18th Dynasty (circa 1600 BC). More recent archaeological and genetic studies suggest domestication was under way before these dates.
A small number of F chaus skulls have been recovered from Egyptian burial sites, but no skulls with features intermediate between F s lybica and F chaus have yet been reported. The Egyptians kept and mummified a number of wild animals; the presence of F chaus bones does not indicate it was breeding with domestic cats or that it was kept as a pet. Some Egyptian paintings depict cats with features consistent with F chaus in a marshland setting.
The role of cats in Ancient Egyptian religion is well documented so there is no need to describe it in detail here. Cats were kept (and bred) in temples where they would have been in close proximity with humans and dependent upon them for food. This would have contributed to their domestication (or their continued domestication) as the tamest cats would have thrived best in such closed and continued proximity with people. Numerous cat mummies have been found; many appear to have been sacrifices and cats would have been reared in large numbers for this purpose. Because of their religious status, as well as their rat-catching abilities, cats were also kept by Egyptian households.
Paintings suggest cats were used in hunting of wildfowl although this is debatable since cats are not generally good retrievers. Later they were depicted in paintings as symbols of fertility and/or domestic harmony.
Domestication of Cats in Ancient Pakistan
Ancient sites have been discovered in the Indus Valley (now Pakistan) dating back to 6000 BC. The Indus had a flourishing civilization with several major cities comprising 40,000 to 50, 000 inhabitant at the time of the Egyptian kingdoms. Domestication of cats may have followed a parallel route to that known to have occurred in Ancient Egypt. Genetic studies do not support independent domestication of the cat. It appears domestic cats reached Asia relatively early along trade routes between ancient empires.
Indus valley sites (e.g. Harrapa, Mohenja-daro) indicate the presence of granaries and the domestication of animals (Jarrig & Meadow, Scientific American 243:122-133, 1980) such as cats, dogs, goats, sheep and possibly elephants. At its peak, this civilization stretched across the whole of Sindh, Baluchistan, Punjab, Northern Rajasthan, Kathiawar and Gujarat. The cities were far more advanced than their counterparts in prehistoric Egypt, Mesopotamia or elsewhere in Western Asia. In common with most other contemporary civilizations, agriculture was the backbone of the Indus economy and the granaries would have attracted rodents which in turn attracted cats.
Bones of F catus were found at some sites in the Indus valley in situations which indicate domestication. There is evidence of cat tracks in mud-and-straw bricks used at these sites (much like cat tracks from where cats walk across drying cement). Whether the F catus bones date back as far as 6000 BC wasn't reported. Studies of Asian sites dating back to 5000 BC shows the domestication of animals in this area well before 3000 BC. Archaeological studies of villages established on the coast of the Bay of Bengal have also shown cat remains associated with domestication.
Historical and archaeological data and genetic evidence (genetic divergence between populations), suggests an origin for the domestic cats of Pakistan in western Asia that predates the Harrapan civilization. The similarities between the Indus and Ancient Egyptian civilizations gave rise to a theory of a west-central Asian centre for cat domestication independent of Egypt (M Ahmad, B Blumberg, M. F Chudary, Journal of Heredity 71:323-330, 1980). Later studies indicate a single domestication event in the Fertile Crescent from which domestic cats spread with traders and settlers.
The Indus cities had commercial links with Afghanistan, Persia, Egypt, Mesopotamia and the Samaritans. Cats could have travelled between Ancient Egypt and the Indus valley with Phoenician traders sometime around 2000 BC, allowing the two populations to interbreed. After 2000 BC, for unknown reasons, the Mohenjo-daro and Harappan culture slowly declined.
Domestication of Cats in China
According to a study by Yaowu Hu, Songmei Hu, Weilin Wang, Xiaohong Wu, Fiona B. Marshall, Xianglong Chen, Liangliang Hou and Changsui Wang "Earliest evidence for commensal processes of cat domestication" Proceedings of the National Academy of Sciences of the USA (PNAS) December 16, 2013 Felis bieti, sometimes considered a subspecies of F lybica, and extant in China, appered to have self-domesticated in order to take advantage of prey species attracted to human homes and farms. Chinese cat fanciers has insisted that the Dragon Li breed was primarily descended from the self-domesticating F bieti, but this appears to have been a marketing ploy. A study in the Proceedings of the National Academy of Sciences of the USA noted that 8 feline bones found in an ancient Chinese village suggested that the advent of farming – the foundation of human civilization – was bound to attract wildcats, which would ultimately self-domesticate as is known to have happened in Egypt and in the Indus Valley.
The study investigated a 5,000-year-old village known as Quanhucun in Shaanxi, central China. The villagers were millet farmers. The archaeologists unearthed rodent burrows tunnelled into grain storage pits and V-shaped ceramic vessels apparently designed to protect stored grain from vermin. In addition, the villagers seemed to have welcomed cats as the feline bones discovered at the site all contained specific signatures of carbon and nitrogen that suggest the cats ate grain-eating rodents. In return, the villagers appear to have cared for the cats. A feline jawbone found at the site showed significant wear on its teeth, indicating a cat that was 6+ years old - elderly by the standards of that time - and had survived to that age because of human care. Analysis of a bone belonging to a different cat indicates that millet-based human food formed a large part of its diet, perhaps due to being a pet fed on human food. The Chinese cats were outside the wild range of Near Eastern wildcats and were biometrically smaller than the Near Eastern Wildcat, but they were within the size-range of domestic cats.
While dogs became hunting companions, at least some of the F lybica subspecies of wildcat may have been predisposed to domestication (or at least semi-domestication) wherever human settlements attracted rodents. However, a study in 2025 found that the bones had been misidentified and actually belonged to the Asian Leopard Cat, then extant in China.
Yu Han, Songmei Hu, Ke Liu, Xiao Xu, Sean Doherty, Alexandra Elizabeth Jamieson, Aurelie Manin, Sofia Granja Martins, Miaomiao Yang, Chong Yu, Juan Wang, Zhuang Wu, Canping Chen, Sicheng Han, Daowei Lu, Lanhui Peng, Xianzhu Wu, Ziyi Li, Wenquan Fan, Quanfa Cai, Zongliang Cui, Jing Yuan, Zihan Li, Yang Liu, Zhipeng Li, Zhendong Liu, Qian Ma, Jing Shao, Zhouyong Sun, Fulai Xing, Wuzhan Yang, Shugang Yang, Lianjian Yue, Pengcheng Zhang, Weilin Wang, Huanyuan Zhang, Yan Zhuang, Xin Sun, Yan Pan, Xiaohong Wu, Laurent A. F. Frantz, He Yu, Joel M. Alves, Greger Larson, Shu-Jin Luo. Leopard Cats Occupied Human Settlements in China for 3,500 years before the Arrival of Domestic cats in 600-900 CE around the Tang Dynasty. bioRxiv, doi: https://doi.org/10.1101/2025.01.31.635809
Yu Han et al analyzed 22 feline bones from 14 sites across China spanning 5,000 years. The earliest cats in human settlements in China were not domestic cats (Felis catus), but native leopard cats (Prionailurus bengalensis). Leopard cats began occupying human settlements around 5,400 years ago up until 150 CE. The remains had previously been misidentified as those of domestic cats. The leopard cat appears to have disappeared from human settlements when the climate became colder and dryer and agriculture declined; the niche it had occupied vanished. There were no archaeological cat remains in China for around 600 years, until the first known domestic cat (706–883 CE) was identified in Shaanxi during the Tang Dynasty. Genomic analysis suggested a white or partially white coat (and underlying pattern mackerel tabby) and a link to a domestic cats from Kazakhstan, indicating a likely dispersal route via the Silk Road. This result indicates a relatively recent arrival of domestic cats in eastern Eurasia, challenging the commonly held assumption that domestic cats were present in China as early as the Han Dynasty (206 BCE–220 CE).
Before domestic cats arrived, all but one of the feline remains found in Chinese archaeological sites have now been identified as leopard cats (P. bengalensis). The only exception was an Asiatic wildcat (F. ornata) found at Huoshiliang.
Whereas there has been a great degree of hybridization between domestic cats and European wildcat species, the Chinese study found no o evidence of widespread hybridization in ancient China between domestic cats and local wildcat species (F. ornata [Asiatic wildcat] and F. bieti [Chinese mountain cat]). Domestic cats relatively recently reached the Qinghai-Tibetan Plateau region where they encountered F. bieti. Recent genetic analysis indicated that modern domestic cats in the Qinghai-Tibetan Plateau region now possess 1-10% ancestry from Chinese mountain cats (Felis bieti). There are concerns about gene flow in the opposite direction, affecting the genetic purity of Chinese mountain cats.
Domestication of Cat Species Elsewhere
The first human cities appeared around 5000 BC (Catal Huyuk, Turkey). Civilization began much earlier in warm climes (Egypt, Indus valley, South America, Mediterranean) than in northern parts of Europe which were still recovering from a cold period. This put the tractable species of wildcat into close proximity with man, especially where they hunted rats attracted to city granaries. It is conceivable that cat species were tamed in other places where humans built cities.
The Mohics of Peru were known to worship the cat as a god of copulation and fertility. It is possible that small species of cat were tamed and kept as pets or in temples. The Jaguarundi is believed to have been kept as a pet although it was tamed rather than domesticated.
Cheetah have been used for hunting, but they were tamed and trained rather than domesticated. Servals have also been tamed and kept as pets. The larger species (Lions, Leopards, Puma) have been kept as pets, but (barring a few exceptional individuals) have an unreliable temperament in captivity and their large size made them dangerous. A few people have kept semi-tame Scottish Wildcats (F silvestris grampia), but these generally revert to wildness as they mature as do modern F s grampia x F catus hybrids.
Domestication of Cats in the Americas
The domestic cat was taken to the Americas by European settlers. Some were ship's ratters, polydactyls being preferred for this task which accounts for the high incidence of polydactyly on the eastern coast of the United States (i.e. the original colonies) and a few eastern coastal locations in Canada corresponding to similar polydactyl populations in south-western coastal England. Others were pets which accompanied settlers. It is possible that cats accompanied the original Viking settlers, however no evidence of feline occupation remains.
There have been reports of early colonists finding domestic cats already in North America and it was, for a while, supposed that the cat may have interbred with native lynx. The size difference between the two species is great enough to prevent natural interbreeding and the greyish creatures with banded tails reported by early naturalists were probably racoons.
In South America there are two cat species which show a high degree of tameability in captivity: Geoffroy's Cat and the Margay. Both have recently been hybridized with domestic cats; the Margay to produce the "Bristol" (later incorporated into the Bengal breed) and the Geoffroy's Cat to produce the Safari. This has caused some naturalists to wonder why these two species were apparently never domesticated.
The most likely reason is that they are forest cats and that forest peoples did not form static societies like those in Egypt and Pakistan. They were primarily hunter-gathers and did not build granaries and therefore had no use for ratters to protect their grain. There are reports that Jaguarundi kittens were occasionally tamed as pets, but there was no concerted effort to domesticate the Jaguarundi and in any case, the Jaguarundi tended to attack domestic fowl.
The small cats in regions where Central and South American peoples did build cities were probably not amenable to domestication. Hence no small cat species was domesticated in the Americas.
The Spread of the Cat to Europe and America
Although Ancient Egyptians forbade the export of cats, by 1700 BC the cat was depicted in domestic poses in Israel (recent studies suggest the cats were already established in those area before the Egyptian prohibition). By 1400 BC domestic cats were present in Greece. By 1000 BC cats had travelled northwards across the Mediterranean aboard ships (possibly with Phoenician traders) and from there they spread along trade routes. The cat travelled eastward to China and Japan (where it protected silkworm cocoons from rats). The Romans regarded cats as rare and exotic pets, preferring the mongoose for vermin control. By 500 BC, domestic cats seem to have been familiar in southern Europe.
The cat may have arrived in England with the Phoenicians who traded for tin in Cornwall, though it is most likely the Romans who first brought cats with them some time before 4 AD. Cat footprints have been discovered in clay tiles at Caesaromagus (Chelmsford, Essex). Further cats arrived with the Vikings.
Oswald Jarrett, former Professor of Comparative Virology at the University of Glasgow Veterinary School, researched methods to control feline leukaemia virus, feline immunodeficiency virus and feline infectious peritonitis (feline enteric coronavirus). Professor Jarrett has found that the subtypes of FIV in Italy and Great Britain are quite different, based on sequencing about 50 isolates in each country. One reason for this striking difference may be that cats and FIV came to Italy and to Britain by different routes from the Near East. The obvious routes are the two by which farming spread, either the southern route along the Mediterranean Sea or the northern route up the Danube (and possibly other rivers out of the Black Sea) through the Hungarian plain into northern Europe. Did cats follow farming? The evidence from sequencing a small number of FIV isolates from other parts of Europe is consistent with this distribution. The distribution of the viruses adds very useful information about both the spread of cats and farming practices in Europe.
While cereal crops apparently reached northern Europe by 5000BC, it is believed that domestic cats reached Britain some 3000 years later, around 2000BC. It is possible that cats were not required by farmers for vermin control until then. While the Romans stored grain in granaries, it had previously been stored in underground pits that might have been less prone to pests - or perhaps the pests themselves had not yet arrived. The spread of cats throughout Europe might be more closely linked to the spread of agricultural pests rather than to cereal farming itself. The house mouse, originally native to parts of Asia, spread to the Mediterranean Basin about 8000 BC and to the rest of Europe around 1000 BC, exploiting human settlements. The Black Rat was established in the Near East in Roman times (though it didn't arrive in Europe until the 6th Century). It seems likely the Romans brought along cats because rats and mice were recognised pests back home and they anticipated similar pests being present in Britain. Domesticated predators, that could be controlled, were preferred to native wild predators (which were often persecuted as perceived threats to poultry). This might account for the lag between cereal farming and the spread of cats.
From then on, we know that cats travelled alongside man in ships, valued as rodent controllers who protected ships' supplies and as lucky mascots. They travelled to America with colonists; the preference for polydactyl cats as lucky shipboard mascots is reflected in the high numbers of polydactyls in the first settled areas on the eastern US coast. It is generally considered that the domestic cat migrated into the Americas alongside European explorers and settlers following the voyages of Columbus. By the time cats came to live alongside humans, there were no land bridges from Northern Europe to the New World. This is supported by the fact that domestic cats don't migrate over long distances of their own accord - they travel as part of human households alongside other possessions in carts or boats.
Ruiz-Garcia (Ruiz-Garcia, Alvarez 2008) found that Latin American cats shared genetic ancestry with Spanish cats, indicating that cats had reached Central and South America aboard Spanish ships. Columbus's ships carried cats. His first two westward voyages ended up at Caribbean islands. His third voyage (1498) reached the Venezuelan mainland and his fourth (1502) reached Central America. Spanish cats may have been taken ashore at any of those places. Throughout the 1500s, Spanish explorers established colonies in South and Central America and would very likely have taken familiar animals with them. By 1537 cats may have reached inland to Paraguay, the centre of South America, with Juan de Salazar de Espinosa.
Discounting (for the time being) a short-lived Viking settlement in Canada, the first Europeans to reach North America were probably Spanish slave traders (based in the Bahamas) in the 1490s, followed by the Portuguese in 1502 (Florida is depicted on the Cantino map). Juan Ponce de Le n is credited as the European discoverer of Florida in 1513. Cordoba visited Florida in 1517 and Cortes arrived there in 1519. Spanish/Portuguese cats may have arrived with any of these. Having probably disembarked in the Americas in the late 1400s and early 1500s, humans and cats would intially have spread up and down the coast, and inland via river trade. Until the early 1600s Spain and Portugal were the most significant colonial powers and would have taken cats from the Iberian Peninsula.
There is some archaeological evidence that the Vikings briefly settled on the American North Atlantic coastline around 1000 AD and it has been suggested that domestic cats accompanied them. Around 995, Vikings established a short-lived settlement at Vinland (Newfoundland). War between those colonists and the native people ("Skraelings") meant they left after only 3 years. There is no archeological evidence at l'Anse aux Meadows of cats surviving after the Vikings departed. The environment is hostile and foxes and coyote routinely hunt cats.
Count de Buffon's Natural History (1767) suggests wild cats existed in pre-colonial America and that a hunter brought a wild cat to Christopher Columbus. The "cat" was described as brown/grey creature, of normal size for a domestic cat and with a long tail. It has been suggested that the cat was either a European domestic cat and that these had survived unchanged over the centuries, or that it was a domestic x F rufus hybrid. The colouration is consistent with either a Bobcat (or hybrid) or an Abyssinian-type cat. However, the term "cat" has been used over the centuries to describe cat-like non-felids (e.g. Pine Martens were once known as Tree Cats) so the creature described might well have been a non-felid, probably a raccoon. The absence of domestic cat bones among the archaeological evidence suggests domestic cats did not reach America until the first European settlers took them there.
Adalsteinsson and Blumenberg suggested that some cats from the north-eastern USA have genes suggestive of a Scandinavian origin. Genetic tests in the 1980s were relatively unrefined and even the researchers admitted that their paper was "idiosyncratic"! The DNA samples were from a variety of cats in several New England cities, especially Boston (which has had extensive sea-trade with Europe). Cats from Northern Europe would have arrived with the original British colonists; with the French whose colonies stretched from eastern Canada to New Orleans; the Swedes (along the Delaware river), and the Dutch (around Delaware & New York). Germans also arrived in the form of the "Pennsylvania Dutch". The Norse or Scandinavian genes found by Adalsteinsson and Blumenberg most likely came from these north European sources, or from early fancy cats traded in the late 1800s. Longhaired fancy cats were partly derived from "Russian Longhairs" while blue "Archangel" cats contributed to the gene pool of early Russian Blues and "Maltese" cats ("Maltese" cats were very popular in the USA). More recent studies found North American cats share genetic ancestry with cats from Finland. The researchers were not versed in cat fancy history (correspondence reveals they'd never heard of the Norwegian Forest Cat breed).
Generally, the cats that spread to Latin America would have been from Iberian stock, while those colonising other parts of North America were of northern European stock. Over the centuries, these populations would have met up as the colonists expanded.
The cat officially penetrated Australia with Europeans in 1788, though aboriginal histories indicate that cats were already present on parts of the Australian coast as shipwreck survivors. Some cats purportedly had Oriental type, suggesting their ancestors had come from Thailand and the Malay peninsula. There are no indigenous species of feline in Australia.
From Notes and Queries Vol 11, 2nd Series (274), Mar 30 1861: In answer to Mr Keightley s observation that cats were as plentiful on the shores of the Mediterranean as in England, I would reply that in Dr Shaw s Travels in Barbary, in the enumeration given of the animals, wild and domestic, of that country, there are no domestic cats. Monsieur Jean Barbot, as late as 1732, testifies to the enormous amount of rats in West Africa, and the comparative scarcity of cats, whose breed the natives distinctly stated came from Europe. From what part of Europe, and at what date let the tale of Whittington of England and Alphonso of Portugal supply the answer. There is no doubt, from Alonzo de Ovalle s Narrative of Chili, that the first imported cats brought great prices in the countries where they had been previously unknown; hence probably the parallel instances of a similar tale to which Mr Keightley alludes: for if the first exported cat produced to its owner so good a price, he would unquestionably, have many imitators, and cats would, for some few voyages at least, be no bad article of commercial speculation. - Samuel Lysons. ( Notes and Queries was a weekly periodical in which scholars and interested amateurs exchanged knowledge on many topics.)
Domesticability of Small Cat Species
The following summarises successes and partial successes in taming other small cat species.
Although normally secretive and shy, the African Wildcat (F s lybica) often lives close to man. Kittens are easily reared in captivity and become docile. When describing the region south of the Bahr el Ghazal, explorer George Schweinfurth wrote: "One of the commonest animals hereabouts was the wildcat of the steppes. Although the natives do not breed them as domestic animals, yet they catch them separately when they are quite young and find no difficulties in reconciling them to a life about their huts and enclosures, where they grow up and wage their natural warfare against the rats. I procured several of these cats, which, after they had been tied up for several days, seemed to lose a considerable measure of their ferocity and to adapt themselves to an indoor existence so as to approach in many ways the habits of the common cat." This is unsurprising; the African Wildcat is the wild ancestor of the domestic cat.
A manul or Pallas's Cat (F manul) from Ladak kept by Colonel A E Ward (1925) became very tame, but disliked strangers. There have been reports of this cat being kept in a semi-domestic state in various parts of Central Asia. A Leopard cat obtained as a kitten by Clifford Pope of the American Museum of Natural History became very friendly and followed Pope about like a puppy. Dr Hans Petzsch of the Halle Zoo found the leopard cats there became as tame as domestic cats. Blyth reported the Fishing cat to be particularly tameable and kept several in captivity. His only mishap was when a newly caught male Fishing cat broke into a neighbouring cage and killed a female leopard twice its own size.
The Caracal is easily tamed and has been trained for hunting in India and Iran. A brace of them were pitted against each other with wagers being made as to which would kill the greatest number of birds from a flock of tame feeding pigeons. The Bheels of Mhowa used caracals to stalk pea-fowl, kites, crows, cranes and hares. Colonel Ward reported that a Punjabi zemindar kept a small pack of caracals for hunting hares (much like coursing with greyhounds). Servals are said to take well to a life in captivity with some becoming remarkably tame and not at all fearful of strangers entering the house. In spite of this, they have never been kept or trained for sporting purposes in their native Africa..
Temminck's Cat (Asiatic Golden Cat) has been reported as becoming tame in captivity. One bought as a kitten by EP Gee became devoted its master, sitting on his lap, suckling his earlobes and licking his head. It played with domestic cats and with dogs and though it went into the forest to hunt, it returned when called. It came from the Garo Hills and was later given to London Zoo.
The Rusty-Spotted Cat (F rubiginosa) has been kept as a pet. According to Jerdon "I had a kitten brought to me when very young in 1846, and it became quite tame, and was the delight and admiration of all who saw it. Its activity was quite marvellous and it was very playful and elegant in its motions." Robert A Sterndale obtained two of these cats as kittens from a Gond tribesman and found them delightful pets: "They became perfectly tame, so much so that, although for nine months of the year I was out in camp, they never left the tents, although allowed to roam about unconfined. ... At night the little cats were put into a basket, and went on with the spare tents to my next halting place; and on my arrival next morning I would find them frisking about the tent roof between the two canvasses, or scrambling up trees under which we were pitched. Whilst I was at work I usually had one on my lap and the other cuddled behind my back on the chair. One day one of them, which had been exploring the hollows of an old tree, close by, rushed into my tent and fell down in convulsions at my feet. I did everything in my power for the poor creature, but in vain, it died in two or three minutes, having evidently been bitten by a snake. The survivor was inconsolate, refused food, and went mewing all over the place and kept rolling at my feet, rubbing itself against them as though to beg for the restoration of its brother. At last I sent into a village and procured a common kitten, which I put into the basket with the other. There was a great deal of spitting and growling at first, but in time they became great friends, but the village was no match for the forester. It was amusing to see the wild one dart like a squirrel up the walls of the tent onto the roof, the other would try to follow, scramble up a few feet, and then, hanging by its claws, look round piteously before it dropped to the ground." Others have also reported the Rusty Spotted cat to be domesticable (and to interbreed with domestic cats).
Rengger (1830) reported that captive jaguarundis became tame and affectionate, but had to be kept leashed to protect the household's ducks and chickens. Even when leashed they attempted to get at the birds. A jaguarundi that had caught a duckling allowed itself to be picked up by the collar and swung through the air, but did not let go of its prey. When the prey was removed by force, it snapped in fury at its keeper's hand. There are reports of native tribes keeping jaguarundis as pets.
The Feline Diaspora
Before genetics technology was well enough advanced, the spread of cats was based, in part, on coat colours and physical mutations. The mackerel tabby pattern is seen in early depictions of cats and would have provided camouflage. The older the colour mutation, the more time it has had to spread around the world either with natural gene flow between areas or with human intervention. This has given some indication of the spread of cats.
Blue (grey) possibly arose in Asia (Korat) and spread from there into Russia (Russian Blue) and France (Chartreux). The lilac colour may also have appeared in Thailand/Malaysia. Red (ginger) cats arose in Asia but the main spread of this colour seems to be via northern Europe with the Vikings. The blotched tabby mutation is believed to have occurred in Britain and spread throughout the former British Empire with human colonists. The colourpointed pattern arose in Asia and is naturally occurring in Thailand (Siam) and Malaysia, but the main recent spread was via the 19th century British cat fancy.
From historical writings, the longhair mutation either arose in three separate areas (Russia, Persia (Iran) and Turkey), or spread from Russia into Turkey, Persia and surrounding countries, or arose and was developed in Turkey and transported on land and sea trade routes to Europe, the Middle East and Far East. This is based on the supposed origins of the modern Persian breed which was derived from Turkish Angora, Russian Longhair and purported Persian cats. Long hair in domestic cats appears to be an adaptation to cold as the paws do not have long fur underneath; in desert species of wildcat long hair is found on the paws and protects against hot surfaces.
The highest densities of the polydactyl trait was found in Boston, Massachusetts (eastern coat of the USA) with a corresponding high density in ports in south-western Britain indicating the mutation occurred in one of those locales and was spread on sailing ships, thence radiating out by land and sea routes (being favoured by sailors, they are also been found in former British colonies e.g. Malaysia where I found a colony of polydactyl cats near Lake Chini). The kink-tailed trait is most prevalent in Asia and into part of Russia, indicating the movement of cats in that region. The heavy (cobby) body type, once attributed to interbreeding with the European wildcat, was developed in Britain through selective breeding in the 19th Century.
The Lipinski study used a subset of breeds recognised by the Cat Fanciers' Association (CFA) in the USA. The CFA recognises 16 natural breeds derived from regional variants that predate the modern cat fancy. The remaining breeds recognised by the CFA have been developed since the mid 20th Century. In the UK, the birthplace of the organised cat fancy, the GCCF shows a similar split of "natural" and "modern" breeds, however books by Louis Wain and Frances Simpson indicate that some of those "natural" breeds are manufactured breeds resulting from crossing cats from different regions e.g. the Russian Blue was bred with other blue cats such as the Chartreux and British Blue Shorthairs in the 19th century.
While studies of colour mutations provided a rough idea of feline spread, Lipinski et al used microsatellite markers to determine genetic relationships between several cat breeds/populations, their geographical origins and the levels of genetic loss due to inbreeding. Genetic analysis produced clustering patterns showing which breeds were most or least closely related. Random-bred cats from geographical populations fitted into those clusters and determined which regions the breeds were associated with.
In the random-bred cats, genetic analysis showed a split separating Asian and East African cats from all other populations. A second split separated Mediterranean basin populations from the European populations. A third split separated East African cats into a separate and distinct cluster. This resulted in four genetic clusters of cats: Europe, Mediterranean basin, East Africa, and Asia. Cats reached the USA with relatively recent European settlers so the European group may be considered European/American.
Australian/New Zealand cats were not included in the Lipinski study. This is unfortunate because there are conflicting theories over the time of arrival of cats in those regions. Although most cats in Australia are of European origin (arriving on the eastern coast with colonists) some feral cats on the northern coastal areas purportedly have oriental appearance and were anecdotally believed to have arrived with ships from the Malay peninsula several centuries before European settlers. Genetic analysis might have resolved whether feral cats on Northern and Eastern Australian coasts had different ancestry.
Asian random-bred populations aligned with the Asian breeds and these were distinct from other populations. This was further split into a Southeast Asian cluster, a Western European cluster, and a cluster comprising all other cats from the Mediterranean basin, East Africa, and Northern Asia. Cats from Singapore (an important shipping port historically and in recent times) were a mix of cats of Asian and European origins.
American random-bred cats grouped with random-bred cats from Western Europe. The two American breeds (Maine Coon and American Shorthair) also clustered with the seven Western European breeds. Domestic cats are not indigenous to the New World and were initially taken there by Western European settlers (accidentally when jumping ship and deliberately as pets and vermin controllers in the colonies). As documented by Frances Simpson, in the late 19th century there was a very close relationship between the American and British cat fancies with breeding stock being exported from the UK to the USA (and their descendents later being exported to the UK). The two American breeds are not significantly genetically different from each other due to their short tenure in that land.
Italian cats appeared to be an admixture of European and Mediterranean stock. Italian and Tunisian cats were a mix of Western European and Mediterranean cats supporting historical ties between Tunisia and Western European countries. Oswald Jarrett, former Professor of Comparative Virology has found that the subtypes of Feline Immunodeficiency Virus in Italy and Great Britain suggest migration of cats to Italy and to Britain/Northern Europe by different routes from the Near East: possibly a southern route along the Mediterranean Sea or a northern route up the Danube (and possibly other rivers out of the Black Sea) through the Hungarian plain into northern Europe.
There was fairly uniform genetic diversity around the Mediterranean region due to frequent movement of ships and caravans on which cats would have travelled. Cats from Sri Lanka and Singapore were a mix of Southeast Asian and European cats. Some of the cats sampled from Singapore were genetically distinct either as a result of earlier British colonialism or recent importation. Sri Lankan cats and the Abyssinian breed were a mix of the East and West. This might reflect maritime trade around the Arabian Sea or British colonialism. The murky history of the Abyssinian indicates hybridization between British cats (the British Tick) and Indian cats.
The Asian cluster of cats was genetically distinct from cats of the Mediterranean basin, Western Europe, and Africa, but there was genetic diversity within the Asian cluster. This indicates they reached the Far East relatively early and then developed in relative isolation whenever trade between successive ancient empires declined. The Asian cluster was further subdivided - populations (purebred and random-bred) from different Asian regions were more genetically dissimilar than different populations within the Mediterranean basin or in Western Europe. Not only were Asian cats isolated from other regions, individual populations of cats within Asia were separate from each other. This may support cat fancy tradition that certain breeds (Siamese, Korat) were perpetuated in Asia very early e.g. as temple cats or palace cats.
The Geographical Origin and Spread of Breeds
Lipinsk et al found Southeast Asian breeds (Burmese (i.e. American Burmese), Korat, Siamese, Singapura, Birman) clustered with the feral populations from Vietnam. Of the Asian breeds, only the Japanese Bobtail grouped strongly with Western cats, albeit with some Asian influence. Cats are not indigenous to Japan, but reached the islands along Asian trade routes hundreds of years ago. The Japanese Bobtail appears in ancient Japanese art and existed back as far as the 6th, but it is claimed that it was introduced into Japan from China at that time, which corresponds to Ida Mellen's account of curly tailed cats in China. The Japanese Bobtail was imported into the USA in 1968 and developed as a breed in the USA. As a result, their gene pool was largely influenced by European/American cats rather than by Asian cats.
The Persian breed unexpectedly (to Lipinski's team, but not to anyone well versed in breed history) was not genetically associated with random-bred cat populations from the Near East, but grouped with random bred cats of Western Europe/America. Historical cat fancy books consider the modern Persian to be derived from Turkish and Russian longhairs crossed with British longhaired cats (Louis Wain, Frances Simpson). It is debateable that original Persian came from Persia. The modern breed was developed from Western European stock and intensively selected for conformation.
The Sokoke, developed from cats found in the Sokoke forest region of Kenya, was closely related to random-bred cats from the Kenyan islands of Lamu and Pate group. The Sokoke was discovered in Kenya and exported to Denmark, thence to the USA. The Kenyan islands and the mainland had shipping ties in the Arabian Sea i.e. to with India, the Near East, the Arabian Peninsula and Africa.
Lipinski et al also analysed the genetic differences within cat populations from Europe/America, East Africa, the Mediterranean and East Asia. The greatest genetic diversity was found in cats from the Mediterranean while Asian cats were less genetically diverse. Several breeds formed indistinguishable pairs: Singapura/Burmese, Havana Brown/Siamese, Korat/Birman (more surprising), and Exotic Shorthair/Persian. Those pairs were distinct from other breeds however. The Burmese showed shared origins with both Siamese and Korats (matches cat fancy history of the breed). The Birman, a colourpoint, might have been expected to show closer kinship with the Siamese than with the Korat.
Three recently derived breeds were not distinct from their parental breeds. The genetic analysis upheld cat fancy history that the Exotic Shorthair is a shorthaired variant of the Persian. It upheld tradition that the Siamese is the ancestor of other Colourpoints along with Orientals and Havana Browns (the latter is considered a chestnut Oriental outside of the USA and not as a separate breed); although recognised as different breeds by cat registries, these differed by as little as one gene variant e.g. for hair length, colour or pattern. The Burmese and Singapura grouping was another example of recent breed derivation and supported claims that the Singapura was derived from Burmese cats exported to Singapore and the descendents reimported to the USA (Burmese cats identical to Singapuras in colour and similar in conformation have been bred in Europe).
The shared origins of Burmese, Siamese and Korat is not surprising. The Burmese derived from a female (Wong Mau) intermediate in type between Siamese and Burmese (this naturally occurring type is now recognised as the Tonkinese). There are naturally occurring intermediates between Siamese and Korat in the form of colourpointed Korats. The close relationship of Korat and Birman is more surprising; there are several versions of Birman history ranging from temple cat in Burma to wholly developed in France.
Persians and Siberians showed within-breed genetic differences. Historical cat fancy books show the Persian was derived from a broad genetic base (Turkish and Russian longhairs crossed with British longhaired cats) but has undergone intense selection over 100+ years. Some Persians also have Siamese ancestry due to interbreeding with the Himalayan/Colourpoint Persian which was derived from Persian/Siamese crosses in the early 20th Century (in the USA, Colourpoint Persians are recognised as a separate breed).
The Siberian, a natural breed and the most recently recognised CFA breed at the time of the study, was more genetically diverse than other purebreds in the sample. It was as genetically diverse as random-bred populations. This indicates that it was derived from a broad foundation stock.
The Sphynx, which Lipinski et al described as a derivative of the Devon Rex breed, also had high genetic diversity. In fact the Sphynx gene pool was expanded using another spontaneous hairless mutations found in the USA (possibly overlooked by the researchers) and these have been crossed with the Devon Rex.
More surprisingly, the Turkish Angora and the Turkish Van were distinct from each other, the Turkish Van being closer to Egyptian random-bred cats while the Turkish Angora was closer to random-bred cats from Tunisia and Turkey and to the Egyptian Mau. Cats may have spread into different regions of Turkey from different directions: into the Lake Van region from Egypt via land trade routes and into the Ankara (Angora) region from Tunisia via sea routes. Cat fancy history has referred to "Russian Angoras" suggesting a link between Russian and Turkish longhaired cats.
However, the UC Davis Turkish Cat Studies of 2008 and 2012 only studied American cat fancy registered Angoras rather than the true Turkish Angora or Ankara Kedisi directly from Turkey, and especially from the Ankara Zoo. The American cats were tested based on cat fancy pedigrees and assumptions as to their ancestral origins in Turkey. Samples submitted from Turkey and the Ankara Zoo were allegedly ignored. Samples claimed to be from Turkey which show a genetic relationship to American Angoras cannot be verified (some named clinics now deny having provided samples). Some were allegedly submitted in the name of vet clinics that had been closed years previously. Independent DNA studies were carried out using documented Turkish Angoras from the Ankara Zoo, Ankara City, from other parts of Turkey, and also Cyprus. The results do not correspond well with that of the American Angora resulting in an erroneous conclusion that the (American-bred) Turkish Angora was most representative of cats of Turkey. Breeders in Turkey feel that the cat fancy s fine-boned version of their national breed is unrepresentative of the true Turkish cats, which are much sturdier. American Turkish Angoras may have only a minimal remnant of the original Ankara Zoo DNA and are only purebred on paper.
The British Shorthair, Exotic Shorthair, Norwegian Forest Cat, Persian, and Siberian each showed subdivisions within each breed, indicating multiple lineages. This is not surprising in the Norwegian and Siberian as these come from a broad base of random-bred cats that have relatively recently been recognised as a breed. The Persian was developed from a range of longhaired cats in the 19th Century (and this diversity is inherited by the derivative Exotic Shorthair). The British Shorthair was developed from random-bred shorthairs in the 19th Century and, for part of that time, the Russian Blue was bred with British Blue Shorthairs before being considered a separate breed..
Attempts to Classify Breeds
At the beginning of the cat fancy, breeds were classified as long-haired (i.e. Persian & Angora types) , short-haired (British Shorthair type) and Foreign (everything else) and as more breeds were introduced, they were fitted into these categories. The following is an alternative classification suggested by Isabella Cole and based on naturalists' observations rather than cat fancy traditions. Some of the variety names of the Asiatic cats are unfamiliar, but there is recognition that that American Siamese and American Burmese differ from the Asiatic versions.
The Cat Gazette: The Cat in America by Isabella Cole (All-Pets Magazine, March, 1946)
Glover M. Allen in "Mammals of Mongolia," 2 volumes, 1938, divides the small group cats into two types:
1. Small cat, tiger typed, tail length of body and similar to Felis Sylvestris and Felis libyca.
2. Short tailed, lynx type, cat, Felis chaus and its allies - of which one race seems to extend to western borders of China. George Mivert in The Cat, 1881, mentions numerous smaller, typed cats with bobtails that inhabit the Malayan peninsula and the East Indies. His details are scanty and his information appears to have been taken from the measurements of skins deposited in the British Museum rather than from first hand information, like the more recent authors. From these researches it may be concluded that two types of domestic cat, based upon structural differences, are recognized today, the tiger type and the lynx type.
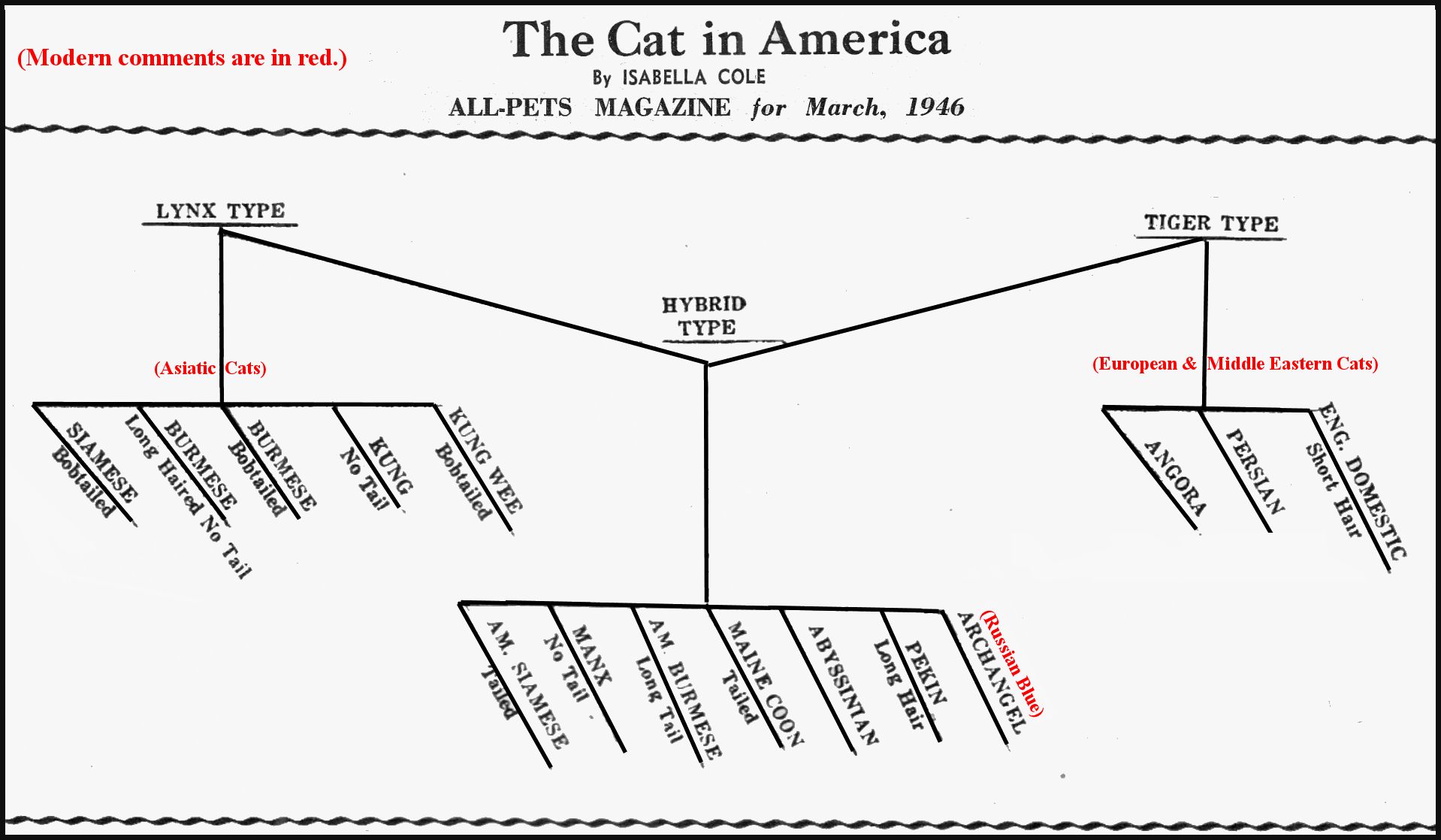
Would it not be more scientific to classify the breeds accepted by the cat fancy into these two types, rather than divide them by color and length of fur? A mixed or hybrid type would be given an intermediate class. The tiger type would include all cats with round heads, domed foreheads, small ears, short nose and foreface, eyes round and not slanting to the nose, body long and slender, back straight, and belly line straight with no roach or elevation of rear quarters, tail the length of body, legs of the same length. Fur can be short or long and any color.
Lynx type would include wedge shaped heads, flat foreheads with no nose stop, or dent, a long nose and a very strong jaw, very long pointed ears set high on upper comers of triangle and held upright, eyes slanting 45 degrees toward the nose, whiskers turned downward and eyebrows curved forward and abundant, body short, compact and very hard and muscular with heavy bone, body much heavier at hips like kangaroo and civet, flank very deep, producing a downward slant toward rear of belly line, hips roached like an English bull dog, rear legs longer than forelegs, feet large and muscular, tail short or tailless, fur short or long and any color, often double and open.
The hybrid type would have a mixed type of conformation and permit a long list of new combinations for new breeds.
The Genetics of Domestication
While European and Asian wildcats and the domestic cats are similar in size, 9,500 years of domestication has made them very different in temperament. Around 9500 ago, wildcats exploited the rodents attracted to grain stores in the Middle East. The friendliest of those cats began to exploit humans for food scraps and shelter. Fed and protected, those cats evidently had greater breeding success so they passed on the friendliness traits and domesticated themselves. Over several thousand years, domesticated cats became slightly smaller and developed various colour and pattern mutations. They retained more of their juvenile behaviour and became more-social with their own species (a sort of arrested development!). The domestic cat s brain is slightly smaller, as are its adrenal glands, and its gut appears longer in order than its wild cousins in order to increase digestion times for a less carnivorous diet (i.e. a proportion of non-meat scraps). An increased degree of white spotting is another characteristic found in domestic animals including cats and was seen as a side-effect in Belyaev s selectively-bred domesticated foxes.
In 2014, a research team led by Michael Montague, of the Washington University School of Medicine in St. Louis, and including William Murphy, a geneticist at Texas A&M University, College Station identified some of the genes responsible for these changes. Their analysis was published in the Proceedings of the National Academy of Sciences in November 2014. They began with a comprehensive feline genome derived from 22 domestic cats. They identified genes (different genes are given gene ID numbers) and compared the resulting genome with those of cows, tigers, dogs, and humans. In cats, the researchers identified 281 genes that showed signs of recent, rapid and/or frequent mutation; these were changes that would have occurred during domestication. Some affected hearing and vision (the cats most important senses) while others played a role in fat metabolism and appear linked to the cat s obligate carnivore lifestyle.
When the researchers compared the domestic cat genomes with that of two Near Eastern wildcats and two European wildcats, they found at least 13 genes that changed as cats transitioned from wild to domestic. These 13 genes were known to affect cognition and behaviour, including the fear response and the ability to learn new behaviours in return for food. During self-domestication, cats became less fearful of new situations and sociable with humans (and other cats) in return for food. Another gene coded for glutamate receptors which play a key role in learning and memory and may have allowed the brains of domesticated animals to develop in such a way that they became more social. The team identified five genes in domestic cats that influence the migration of neural crest cells, stem cells in the developing embryo that affect everything from skull shape to coat colour. These cells may be the master control switches of domestication, explaining why domestic animals share common traits, such as smaller brains and certain pigmentation patterns (e.g. white spotting), something noted by Charles Darwin.
The cat genome appears to have undergone less intense and more recent evolutionary pressure than the dog genome. Dogs have lived around humans for 30,000-ish years and the various breeds were developed for specific purposes. Cats, however, exploited humans from a much later date and were only intensely developed into fancy breeds from the 19th century (excepting those natural breeds that arose due to mutation in a geographical area e.g. Angora longhair, Siamese pattern). While housecats are thoroughly domesticated, they have kept their options open and most retain enough wild instincts to revert to the wild state as feral cats.
Effect of Selective Breeding on Genetic Diversity
As well as the relationship between breeds, researchers found artificial selection had reduced diversity within single breeds. This is due to breeders striving for a consistent look and limiting the breeding population. Studies confirm that cat breeds are less genetically diverse than random-bred cats. Pure breeding is generally associated with a loss of genetic diversity. In the breeds sampled by Lipinski et al, the loss of diversity did not correlate with breed popularity or age, but did correlate with the number of foundation animals from which the breed was established and the allowable outcrossing to other breeds. The Burmese, Havana Brown, Singapura, and Sokoke were least diverse.
The Burmese and Singapura breeds were the least genetically diverse of the breeds sampled. This reflects the most intense inbreeding . The Burmese is descended from a female called Wong Mau (who is now considered a Tonkinese). The Singapura is largely a derivative of the Burmese although there is a fanciful mythology about it being an indigenous Singaporean cat (supporting this, in Holland there are cats identical to the Singapura that have been bred from wholly Burmese lines). The Sokoke is a recent breed being developed from a small pool of distinctive looking cats found in the Sokoke forest region of Kenya. The Havana brown is a colour variant of Siamese/Oriental cats.
The Siberian was the most genetically diverse and comparable to the genetic diversity in random-bred cats due to having a broad foundation stock. The Norwegian Forest Cat is likewise genetically diverse.
Unlike dog breeds (some of which are thousands of years old) most cat breeds were developed within the past 150 years, mainly in Europe (particularly Britain where the organised cat fancy originated) and the United States. Around 80 cat breeds are recognised by the major cat fancies. The CFA (USA) recognised only 41 breeds at the time of Lipinski et al's study and it was this list that was used by the researchers. If informal breeds/strains (those that have not achieved cat fancy recognition) are included, there are 200+ breeds around the world, some naturally occurring and some the result of crossing existing breeds. The CFA recognises only a subset of cat breeds.
The oldest CFA recognised breeds are the Persian, Russian Blue, Siamese and Angora (in Britain the Angora and Persian merged while the British Shorthair and is one of the oldest recognised breeds; Turkish Angoras were reimported and re-established in Britain at the end of the 20th century). The study included 15 of the 16 "natural" (foundation) breeds. The one not included was the Manx.
Lipinski et al found the purebreds formed distinct groups. The Southeast Asian breeds included Birman, Burmese, Havana Brown, Korat, Siamese and Singapura. At the other end of the genetic spectrum were the Western breeds. The Abyssinian and the Japanese Bobtails had genetic markers from both of those groups indicating the result of cross-breeding during their development.
The majority of the foundation or modern breeds appeared to have originated from random-bred cats of the purported region of origin (Lipinski et al). The Southeast Asian breeds were strongly associated with the random-bred cats from Vietnam, China, Korea, and Singapore. The Siberian had strong associations with cats from Germany and Finland (samples from random-bred Russian cats were not available to the researchers). (American-bred) Turkish Vans grouped with regional random-bred cats from Turkey, Israel, and Egypt.
Egyptian Maus, which folklore claims originated in Egypt and which resemble pharoanic Egyptian depictions of domestic cats, showed European influences. It appears these cats resemble ancient cats of Egypt rather than having originated from Egyptian cats (or that they have become very mongrelised by outcrossing). It was also closer to Turkey and Tunisia random-bred cats than to Egyptian cats.
As previously noted, the Persian and Japanese Bobtail were both more closely related to Western European/American cats. The Persian ancestry of the Persian breed is anecdotal as previously noted. Bobtailed Japanese cats were imported into the USA where they were refined into the Japanese Bobtail breed through crossing and selective breeding. The Japanese Bobtail was supposedly introduced into Japan from China around the 6th century (Ida Mellen). It would have been interesting to compare the Japanese Bobtail to the indigenous Russian Kuril and Karel bobtails which (visually) are of Western European, rather than Asian, type.
Conclusion
The Egyptian genesis is still the accepted theory and appears confirmed by genetic studies. The discovery of evidence of domestication at Asian sites as early as 4000 BC to 5000 BC opened the question of independent domestication of cats among other ancient peoples. It appears more likely these cats reached Asia along early trade routes. China's ancient archaeology is still relatively unknown and there is the possibility that Chinese species of small cat may have been domesticable.
The fact that cat remains have been found in various ancient cities and settlements in places other than Egypt suggests small cat species have always lived in or near human settlements, probably in a semi-tame state, attracted there by shelter and by food. This made the cat, particularly F s lybica, ripe for domestication. The apparent simultaneous domestication in several distinct regions (with Egypt being the best known example) supports the genetic evidence that domestication was in progress much earlier than originally believed and that domestic cats had already spread to other areas. Early spread seems more likely than multiple simultaneous domestication events.
The involvement of other species in the evolution of the domestic cats is not confirmed by current DNA analysis. Interbreeding with another suggested ancestor, F chaus, appears only to occur where F chaus has no other choice of mate. Interbreeding with F manul is anecdotal or very isolated. Interbreeding with F margarita has not been demonstrated. If such interbreeding has occurred at all during the evolution of the domestic cat, it is so minimal that no gene markers have been found and it can be disregarded when considering the cat's taxonomic status. Bone comparisons showing similarities with wild species other than F s lybica may also be due to independently occurring mutations in genes inherited from the common ancestor of all Felis species.
Footnote 1 - Modern Cats: Wild or Mild?
Is the modern domestic cat still basically a wild animal?
The proliferation of feral cat colonies demonstrates that modern domestic cats retain enough of their wild ancestry to successfully revert to the wild state and to create self-sustaining populations of wholly wild cats, identical to their domestic cousins in every way except for temperament and, over a suitable time period, a return to the wild type as human-selected traits are removed through natural selection. They remain genetically similar enough to F s silvestris/grampia and F s lybica to form fully fertile hybrids with these wild species.
This does not mean that every individual cat can revert to the wild state. Many pedigree cats have been bred into forms unsuited to the wild state - too short noses which prevent hunting, excessive fur which mats and becomes a liability, too little fur so that hypothermia is a hazard, excessive docility or skeletal deformities. Other individuals have had survival traits surgically removed i.e. declawing. In addition, the "wild state" in many areas is full of man-made hazards.
Many behaviours of pet cats are manifestations of their wild instincts e.g. scent marking, hunting toys. Many outdoor-going cats display a full range of wild behaviours e.g. hunting and marking/protecting territory. In indoor cats, instincts frustrated by close confinement can be redirected into problem behaviours which conflict with their attraction as a pet e.g. middening, attacking ankles. It is understandable that humans want to keep pet cats out of harm's way, but unless the captive environment is enriched, the cat acts out its natural instincts as best it can. Some cats retain more wild behaviours than others which is why not all cats are suited to the indoor-only life forced upon them by the human environment.
Whether we like to admit it or not, our domestic cat companions are essentially still wild at heart and in their genes and instincts; something which keeps pet behaviourists in business.
Footnote 2- How Long Does Domestication Take?
How long does it take to domesticate a species? If the species naturally produces some less skittish animals, the answer can be as few as 4 generations. This has been tested in silver foxes, American mink and otters and scientists are beginning to unravel the genetic basis for tameness - which could explain why some species have never been domesticated. This a short summary of some of the research into the domestication process.
Dmitry Belyaev geneticist at the Institute of Cytology and Genetics, Novosibirsk began selectively breeding tame silver foxes in 1959. He obtained 130 relatively friendly silver foxes from a fur farm. In each generation, he allowed only the tamest animals to breed. Within four generations, some of the foxes had started to wag their tails. After eight generations, his tame foxes showed pied colouration. In successive generations, they showed floppy ears, shorter tails, wider skulls and would breed at any time of year. Within 20 years, Belyaev had created dog-like tame foxes that made good pets, as devoted as dogs but as independent as cats.. Physiologically, they had reduced activity of the hypothalamic-pituitary-adrenal axis, the nervous and hormonal signals involved in controlling an animal's response to stress. The tame foxes have higher levels of serotonin in their brains, inhibiting aggressive behaviour.
n 1972, Belyaev began breeding rats - beginning with wild rats and selecting one group for tameness and the other for aggression. After only 30 years of selection, the researchers had a tame group that enjoyed human contact and a viciously aggressive group. Belyaev then acquire 200 relatively tame American mink. After 4 generations of selective breeding he produced mink with clear evidence that domestication was occurring: temperament, new coat colours and other anatomical changes. In 1980, the experiment was repeated with wild river otters. After 13 years and 3 generations later, the percentage of tame otters had risen from 10% to over 30% and the tame otters had lighter markings, changed reproductive cycles and changes in brain chemistry. Belyaev died in 1985, but his work is continued by Lyudmila Trut. When the Societ Union broke up in the early 1990s, the experiments ran out of funding and the foxes and mink were reduced in number while the otter breeding ceased altogether (some tame otters and their descendents are now at Moscow Zoo).
At Max Planck Institute for Evolutionary Anthropology in Leipzig, Germany, Scante P bo and his team are now looking at the genetic changes for tameness. This might help us understand why the horse-like zebra and African Buffalo are intractable and might even explain why some humans behave more aggressively or anti-socially than others. P bo acquired 15 tame and 15 aggressive rats from Novosibirsk to establish colonies in Germany. To check the behaviour was genetic, not nurture, pups from aggressive rats were fostered on the tame mothers and vice versa. However, the pups behaved according to their genetic parents, rather than learning tame or aggressive behaviour from their foster parents. Ruling out whether maternal hormones affected the developing foetus had been done previous by transplanting embryos in tame and domestic silver foxes; the offspring still behaved according to their genetic parents.
The researchers then cross-bred the rats; the second generation of hybrids showed a spectrum of behaviour from very tame to very aggressive. These were assessed for tameness according to whether they could be handled or whether they violently attacked the handler. The results were compared with 200 genetic markers and several key regions of the genome seemed to have a strong effect on tameness. At least 5 genes interacted to produce the overall effect of tameness or aggression. Further study is needed to identify the mutations and exact effects. It might then be possible to replicate these in the genomes of other species that man wants to domesticate, but which have proven intractable so far.
Most domesticated mammals differ from their wild ancestors in terms of body shape, markings and breeding patterns. Darwin believed these changes came slowly and cumulatively. Belyaev suggested humans selected one useful trait - tameness - and took only a few generations to develop a tame population. Human hunter-gatherers might have captured some of the less skittish prey animals and opted to breed them as a reliable food supply kept close to home. Other traits associated with domesticated animals might be byproducts of breeding for tameness as Belyaev found in his research.
Further Reading:
|
MESSYBEAST : SMALL CAT HYBRIDS, BRITISH BIG CATS & PREHISTORY |